植物病害生物防治菌的筛选*
2019-10-16宋瑞清
曹 翠 宋瑞清,2**
(1. 渭南师范学院美术学院,陕西 渭南 714099; 2. 东北林业大学林学院,黑龙江 哈尔滨 150040)
0 引 言
云杉八齿小蠧(I.typographus)属于鞘翅目(Coleoptera)、小蠹虫科(Scolytidae)、齿小蠹属(Ips)[1].主要危害云杉属、冷杉属和松属等植物,猖獗发生时可导致树木成片枯死.小蠹虫伴生菌是指那些由小蠹虫携带、传播到寄主植物上,使寄主植物无法正常生长的一种病原真菌[2].小蠹虫伴生菌是植物-小蠹虫-伴生菌系统中削弱树木抗性、协助小蠹虫侵害的重要因子[3].在小蠹虫-伴生菌的危害体系中,人们倾向于重视小蠹虫的防治,而忽视了伴生菌的防治.
通常情况下,在防治害虫和病原菌的过程中,人们多倾向于使用杀菌剂,而化学农药的大量使用在防治植物病虫害的同时,也杀死了环境中的有益微生物,提高了植物病原菌的抗药性,同时也严重破坏了农业生态系统,造成环境污染.随着生物科技的发展,拮抗真菌在植病生物防治中的应用受到研究者的重视,即用生物方式来防治植物病害,在无人工干预条件下,采用一类或多类生物来减少病原菌数量或病害发生和减弱一些致病的病原体的生存活力,旨在减少由病原菌所导致的灾害.目前已有多种微生物具有生物防治作用,在20世纪30年代,研究显示木霉菌对植物病原菌的拮抗作用[4].自20世纪70年代以来,国内外对木霉菌的拮抗作用及其机制作了许多深入研究,证实了木霉对病原菌的重寄生现象,同时在温室及田间试验中也取得了较好的效果,已经有商品化的木霉制剂问世[5].在防治病害危害植物的前提下应该首先保护植物,文献中,作者强调对植物的保护,同时需要维持生态系统的多样性和平衡性,故利用生物防治病虫害对植物侵害的道路任重而道远[6].因此,本研究以3株云杉八齿小蠹伴生真菌为拮抗目标,通过平板对峙培养法进行拮抗菌的筛选,选择对伴生真菌拮抗效果明显的拮抗菌,为遭受病害的生物提供功能性的菌株.
1 实验材料与方法
1.1 实验材料
研究使用的云杉八齿小蠹伴生真菌和拮抗菌株均由东北林业大学林学院宋瑞清老师实验室提供.其中,伴生真菌包括Ophiostomapenicillatum1(OP1)、Ophiostomapenicillatum2(OP2)、Ophiostomasp(OS).拮抗菌株包括快速生长菌,包括Trichodermacitrinoviride、Trichodermaatroviride、Trichodermaviride和慢速生长菌,包括Alternariatenuissima、Bionectriaochroleuca、Fusariumtricinctum、Gibberellamoniliformis.
1.2 实验方法
采用马铃薯葡萄糖琼脂培养基(PDA)平板对峙培养法进行真菌培养,实验分为实验组和对照组.培养基直径90 mm,相距45 mm分别接入培养6 d、等量的直径1 cm菌落.实验组接入拮抗菌与病原菌菌落,对照组接入各菌株单独培养.菌丝一面向下的菌饼来直接接触培养基.
按照生长速度,将7株拮抗菌株分为快速生长菌株(3株)和慢速生长菌株(4株).培养快速生长菌株时,首先接种伴生菌株,3 d后再接入快速生长菌株;培养慢速生长菌株时,同时接种慢速生长菌株和伴生菌株.每组平行3次,培养温度25℃,每12 h测量菌落的相对生长半径(共4个半径),包括:对峙培养的病原菌和拮抗菌的相向半径以及2种真菌各自单独培养的半径[7].在进行拮抗结果计算时,采用两菌落之间的相向半径进行拮抗生长测定.
被抑制率的定义是拮抗菌和病原菌被对方抑制的抑制率,相对抑制效果的定义为拮抗菌在抑制病原菌时本身同时被抑制消耗,自己被抑制一个单位,即l%时所得到的抑菌效果[8].在同一环境下,真菌群落的各个成员存在着相互的作用,相对抑制效果更加显现拮抗菌的真实作用,1是相对抑制效果的阈值.相对抑菌效果和被抑制率的数学定义如下[7]:
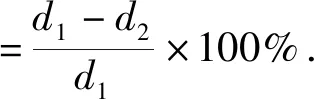
式中,d1为单独培养菌落半径;d2为菌落趋生半径.
相对抑菌效果=病原菌被抑制率/拮抗菌被抑制率.
2 结 果
2.1 拮抗菌株单独培养菌落直径
在相同培养基和相同温度下,不同菌株的生长速度完全不同,菌落直径数据列于表1、2.结果显示,2株云杉八齿小蠹伴生真菌OP1和OP2均在24 h 时菌落已扩展,直径分别为1.15和1.20 cm,伴生真菌OS在24 h后开始生长,在144 h 时,3株伴生真菌菌落直径已达到最大值,分别为8.50、8.43和8.05 cm;4株慢速生长菌(Alternariatenuissima、Bionectriaochroleuca、Fusariumtricinctum和Gibberellamoniliformis)在144 h内菌落以不同速度在生长,在144 h时直径分别达到6.70、3.95、7.45和5.65 cm;3株快速生长菌(Trichodermaitrinoviride、Trichodermaviride、Trichodermaatroviride生长速度快,在48 h时直径均达到最大值8.50 cm.
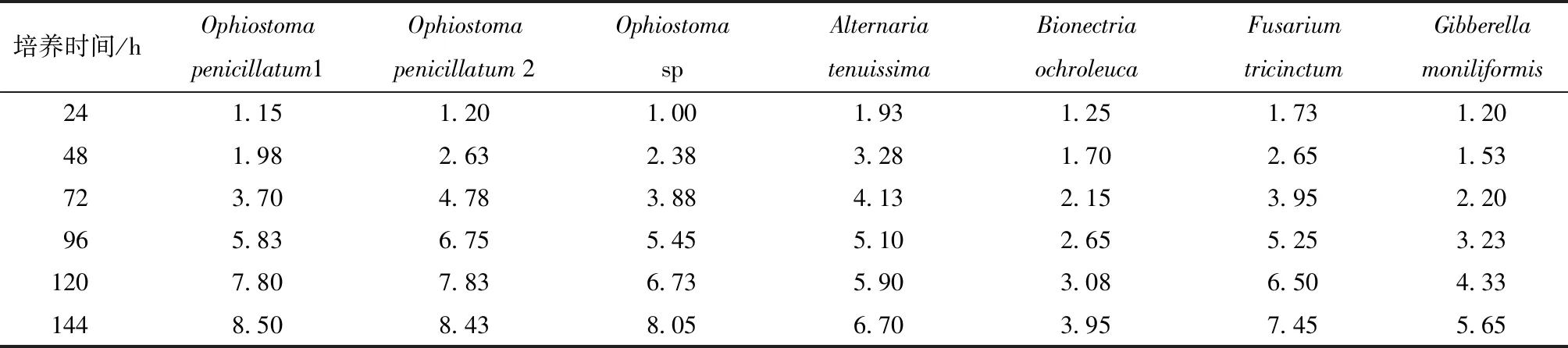
表1 慢速生长菌株单独培养菌落直径/cm

表2 快速生长菌株单独培养菌落直径/cm
2.2 拮抗菌株与OP1菌株拮抗效果
慢生长菌株与OP1菌株的拮抗数据列于表3,相关图像如图1所示.在慢速生长菌与伴生真菌的拮抗过程中,伴生真菌的被抑制率总体呈上升趋势,至72 或96 h开始相互抑制;在Fusariumtricinctum与OP1的对峙培养中,72 h后才产生相互之间抑制作用,各自的抑制率随着时间的增加被逐渐增大,最大值在144 h(55.29%)处;在Alternariatenuissima与OP1的对峙培养中,48 h之前Alternariatenuissima对OP1 的生长有促进作用,72 h后出现抑制作用,随时间的增加,OP1的抑制率在逐渐增大,144 h达到最大(51.76%);在Bionectriaochroleuca与OP1的对峙培养试验中,OP1的生长在72 h之前一直是促进状态,96 h时处于抑制状态;在Gibberellamoniliformis与OP1的对峙培养中,OP1的抑制率随时间的增加而增大,144 h达最高(65.88%).伴生菌株的抑制率明显高于拮抗菌株的抑制率,特别是Fusariumtricinctum在72 h之前,Fusariumtricinctum和OP1处于相互促进生长作用(抑制率为负值);120 h之前Bionectriaochroleuca的生长基本处于不显著的促进状态,之后才逐渐抑制;72 h之前Gibberellamoniliformis生长处于促进状态,96 h后生长抑制.相对抑制效果在144 h时由高至低为:Fusariumtricinctum、Gibberellamoniliformis、Bionectriaochroleuca、Alternariatenuissima,最高值分别达到34.70、6.77、3.98、2.48.综上,对OP1相对抑制效果相对好的菌株为Fusariumtricinctum(34.70).
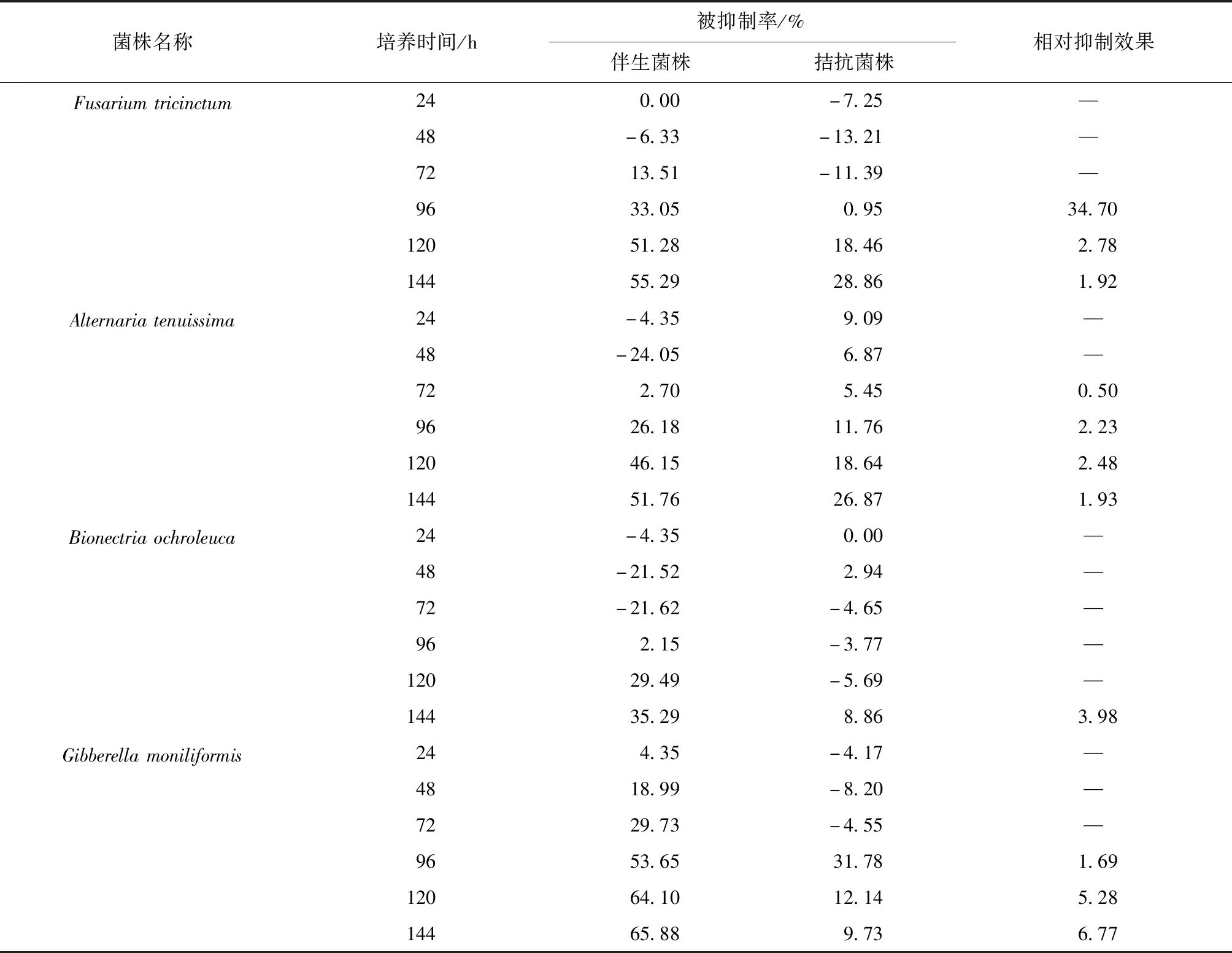
表3 慢生长菌株与OP1菌株的拮抗数据
注: —指无数据

图1 慢生长菌株与OP1菌株的拮抗效果(a)OP1与Fusarium tricinctum对峙;(b)OP1与Alternaria tenuissima对峙;(c)OP1与Bionectria ochroleuca对峙;(d)OP1与Gibberella moniliformis对峙
快速生长菌株与OP1菌株的拮抗数据列于表4,相关图像如图2所示.在快速生长菌Trichodermacitrinoviride、Trichodermaviride和Trichodermaatroviride与OP1的对峙培养中,12 h之前OP1生长处于促进状态,与OP1菌落在36 h时接触,接触以后即出现对峙之势,两菌落停止生长,呈现显著的抑菌带.可能原因是,本研究先接入OP1在培养基中培养,含有充足的营养和具有大量的空间,OP1的抑制率小于快速生长菌.3株生长快速的菌株与OP1之间均出现显著的抑菌带,抗生素在短时间内产生了.在全部对峙培养中,均没有超过1,全部在60 h达到最大(0.29、0.29和0.35).综上所述,表明Trichodermaatroviride对OP1有显著的抑制效果,其他2个菌株有较好的抑制作用.
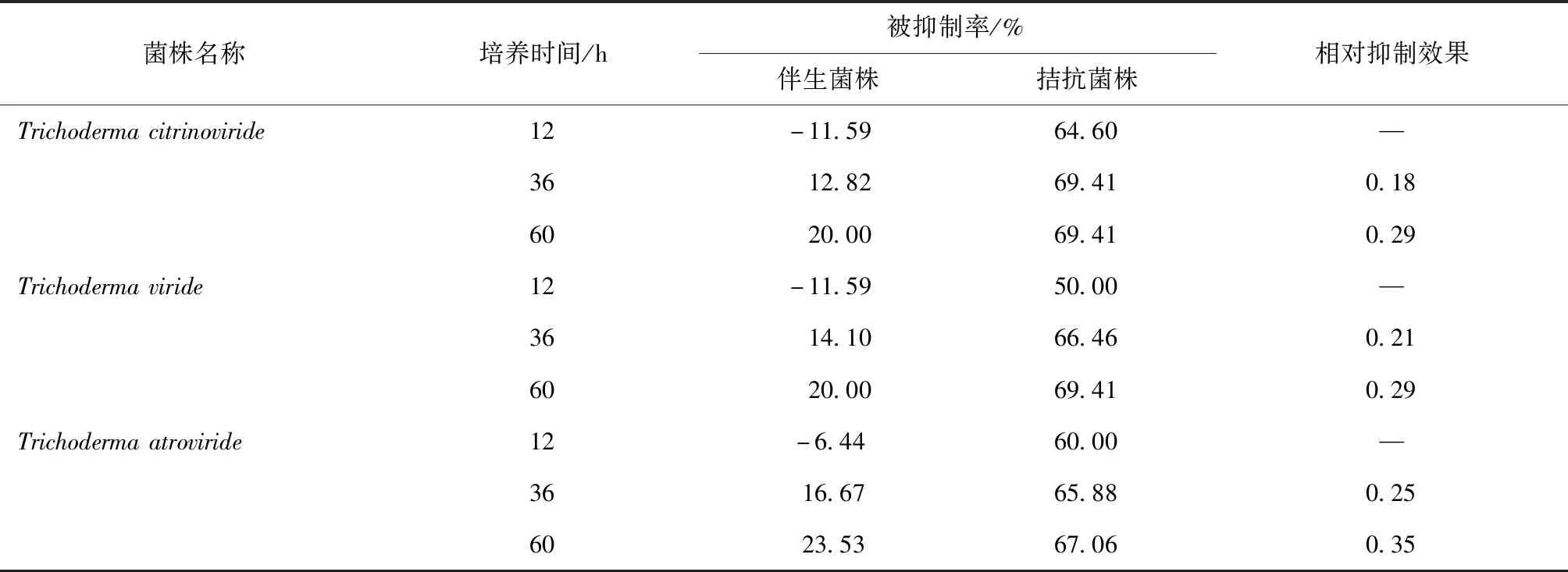
表4 快速生长菌株与OP1菌株的拮抗数据
注: —指无数据

图2 快速生长菌株与OP1菌株的拮抗效果(a)OP1与Trichoderma citrinoviride对峙;(b)OP1与Trichoderma viride对峙;(c)OP1与Trichoderma atroviride对峙
2.3 拮抗菌株与OP2菌株的拮抗效果
慢速生长菌株与OP2菌株的拮抗数据列于表5,相关图像如图3所示.在慢速生长菌与伴生真菌的拮抗过程中,伴生真菌的抑制率总体呈上升趋势,Fusariumtricinctum、Alternariatenuissima和Gibberellamoniliformis与OP2的对峙培养中,随着时间的增加,OP2的抑制率在增大,144 h达最高,抑制率分别为56.08%、51.34%、53.71%;Bionectriaochroleuca与OP2的对峙培养中,24 h之前OP2的生长处于促进状态,48 h后处于抑制状态.拮抗菌株的抑制率明显低于伴生菌株的抑制率,72 h之前伴生真菌OP2对Fusariumtricinctum的生长有显著的促进作用,96 h后才产生抑制作用;24 h之前OP2对Alternariatenuissima的生长有促进作用,48 h后双方生长处于相互抑制状态;120 h之前Bionectriaochroleuca和Gibberellamoniliformis的生长状态一直处于显著的促进状态,之后才逐渐抑制.相对抑制效果在144 h时由高至低为:Alternariatenuissima、Fusariumtricinctum、Bionectriaochroleuca、Gibberellamoniliformis,最高值分别达到33.69、14.78、9.77、8.67.因此,对OP2相对抑制效果好的菌株为Alternariatenuissima和Fusariumtricinctum.

表5 慢速生长菌株与OP2菌株的拮抗数据
注: —指无数据

图3 慢生长菌株与OP2菌株的拮抗效果(a)OP2与Fusarium tricinctum对峙;(b)OP2与Alternaria tenuissima对峙;(c)OP2与Bionectria ochroleuca对峙;(d)OP2与Gibberella moniliformis对峙
快速生长菌株与OP2菌株的拮抗数据列于表6,相关图像如图4所示.在快速生长菌株Trichodermacitrinoviride、Trichodermaviride、和Trichodermaatroviride与OP2的对峙培养中,12 h后产生抑制作用,Trichodermacitrinoviride与OP2接触后即呈现对峙之势,2种菌落停止生长,且出现明显的抑菌带,Trichodermaviride和Trichodermaatroviride与OP2接触后,菌落分别继续生长侵入OP2的菌落而使OP2菌落减小,也呈现显著的抑菌带.被抑制率均随时间的增加而增大,60 h达到最高;OP2首先被接入培养基培养,具有充足的营养和空间,致使OP2的被抑制率小于快速生长菌的.3种生长快速的菌落与OP2的对峙培养均产生明显的抑菌带,表明在短时间内抗生素被产生出来.在全部对峙培养试验中,均没有超过1,全部在60 h达到最大,分别是0.30、0.24和0.35.综上,表明Trichodermaatroviride对OP2有显著的抑制效果,其次Trichodermacitrinoviride也有较好的抑制效果.

表6 快速生长菌株与OP2菌株的拮抗数据

图4 快速生长菌株与OP2菌株的拮抗效果(a)OP2与Trichoderma citrinoviride对峙;(b)OP2与Trichoderma viride对峙;(c)OP2与Trichoderma atroviride对峙
2.4 拮抗菌株与OS菌株的拮抗效果
慢速生长菌株与OS菌株的拮抗数据列于表7,相关图像如图5所示.在慢速生长菌与伴生真菌的拮抗过程中,伴生真菌的被抑制率总体呈上升趋势,至72或96 h开始相互抑制,在Fusariumtricinctum、Alternariatenuissima和Bionectriaochroleuca与OS的对峙培养中,72 h之前处于相互促进中,Alternariatenuissima对OS的生长有显著的促进作用;在96 h后相互之间才产生抑制作用,随时间的增加各自的被抑制率逐渐增大,144 h达最大(分别为46.58%、45.34%和35.4%);在Gibberellamoniliformis与OS的对峙培养中,48 h之前OS的生长处于促进状态,72 h后产生了抑制作用,被抑制率随时间的增加而增大,144 h达到最高(56.52%);拮抗菌株的被抑制率明显低于伴生菌株的被抑制率,在整个对峙培养过程中,OS对Alternariatenuissima和Gibberellamoniliformis的生长一直处于抑制状态,随时间的延长作用一直增大,144 h被抑制率达到最高;OS与Bionectriaochroleuca的对峙培养中,120 h之前Bionectriaochroleuca的生长处于不显著的促进状态,之后逐渐抑制.相对抑制效果在144 h时由高至低为:Gibberellamoniliformis、Bionectriaochroleuca、Fusariumtricinctum、Alternariatenuissima,最高值分别达到4.26、3.11、1.48、1.32.因此,对OS相对抑制效果好的菌株为Gibberellamoniliformis.

表7 慢速生长菌株与OS菌株的拮抗数据
注: —指无数据

图5 慢生长菌株与OS菌株的拮抗效果(a)OS与Fusarium tricinctum对峙;(b)OS与Alternaria tenuissima对峙;(c)OS与Bionectria ochroleuca对峙;(d)OS与Gibberella moniliformis对峙
快速生长菌株与OS菌株的拮抗数据列于表8,相关图像如图6所示.在快速生长菌株Trichodermacitrinoviride、Trichodermaviride和Trichodermaatroviride与OS的对峙培养中,在快速生长菌接入12 h后与OS菌落接触,双方产生抑制作用,接触后快速生长菌菌落分别继续生长侵入OS的菌落而使OS菌落减小.被抑制率均随时间的增加而增大,60 h达到最高;因OS先被接入培养基,具有充足的营养和巨大的空间,致使OS的被抑制率显著小于快速生长菌.各对峙培养均产生明显的抑菌带,说明抗生素在短时间内产生.在全部对峙培养试验中,在36 h时,相对抑制效果大于1,全部在60 h达到最大(分别是2.00、1.47和1.77).因此,对OS相对抑制效果好的快速生长菌株为Trichodermacitrinoviride和Trichodermaatroviride.

表8 快速生长菌株与OS菌株的拮抗数据

图6 快速生长菌株与OS菌株的拮抗效果(a)OS与Trichoderma citrinoviride对峙;(b)OS与Trichoderma viride对峙;(c)OS与Trichoderma atroviride对峙
3 讨 论
在相同的生物环境中,通过采用平板对峙培养法[9],真菌间的相互关系能够很好的反映.本研究通过采用平板对峙培养法,使用相对抑制率和被抑制率两个指标来共同评价7株拮抗菌株对3株伴生真菌的拮抗效果,结果表明,Trichodermaatroviride以其生长较快,能较先抢占营养和生存空间,从而有效抑制了云杉八齿小蠹伴生真菌OP1和OP2的扩展,有显著的抑制效果,Trichodermacitrinoviride对OS同样有显著的抑制效果.慢速生长菌中,筛选出对OP1抑制效果最佳的菌株为Fusariumtricinctum(34.70)、Gibberellamoniliformis(6.77);对OP2抑制效果好的菌株为Alternariatenuissima(33.69)、Fusariumtricinctum(14.78);对OS抑制效果好的菌株为Gibberellamoniliformis(4.26)、Bionectriaochroleuca(3.11);筛选出几种拮抗菌株均具备作为生物防治资源的潜力.
由于接入时间的不同而使快、慢生长菌的相对抑制效果不同.试验结果显示,有部分菌株的生长速度之所以减慢,是由于生长速度迅速的菌株抑制了其生长.于是,Shaw等[10]提出缓慢生长的菌株应该早于快速生长的菌株放入培养皿.由此,二者就可以在培养皿的中心直径处相接.从试验中,可以得出快速生长菌与伴生真菌的各对峙培养均产生明显的抑菌带,说明在短时间内产生了抗生素.快速生长菌在短时间内对伴生真菌产生较强的抑制作用,根据抑制效果的大小来筛选最佳的拮抗菌株.
在使用传统反方法筛选拮抗菌时,是以病原菌和拮抗菌对峙培养时拮抗线的宽窄作为评价的准则.但是在自然界中,显著拮抗的真菌在同一环境中很难同时存在.因此,与室内结果相比,被筛选出的拮抗菌在实际应用环境中,结果并不理想.平板对峙培养可以观察同一生境中的真菌的互相作用,对相互间的影响能够很好的反应[9].本研究所采用的拮抗菌与伴生真菌生活在同一环境中,在此条件下筛选获得的拮抗效果显著的菌种将更适合于实际的应用环境.
