金川荒漠植物光响应曲线最优模型分析
2019-10-16王引弟巨天珍周玉霞万子栋高天鹏杨颖丽
王引弟, 巨天珍, 周玉霞, 万子栋, 高天鹏, 杨颖丽
(1. 西北师范大学 地理与环境科学学院, 兰州 730070; 2. 甘肃省矿区污染治理与生态修复工程研究中心, 兰州 730070; 3. 祁连山北麓矿区生态系统与环境野外科学观测研究站, 金昌 737100; 4. 西北师范大学 生命科学学院, 兰州 730070)
光照强度是影响植物光合作用最明显、最根本的环境因子,其从根本上影响着植物的生长和发育。植物的光合-光响应曲线能反映植物光合作用的强弱,有助于阐明植物生长过程中光合产物的积累与当地环境之间的关系,以及外界环境的变化对植物生长发育产生的影响,对植物光合生理生态学的研究具有重要的意义[1-2]。因此,植物的光合速率与光照强度间的研究显得尤为重要。通过绿色植物的光合-光响应曲线,可直接反映植物的光饱和点以及与绿色植物光合作用所对应的光补偿点、暗呼吸速率、初始量子效率等参数[3],而这一系列的光合参数对研究绿色植物的光合特性具有十分重要的意义,能更加直观地反映不同物种间光化学效率所存在的差异,主要表现在植物生长过程中对光照条件的适应能力[4]、植物体内光合作用机制是否能够正常运转,以及绿色植物在光合作用中利用光强的能力[5]等方面。
光响应曲线作为植物生理生态学研究的重点之一,备受国内外学者的广泛关注[6-11]。诸多学者对已提出的非直角双曲线模型[12]、直角双曲线模型[13]、指数模型[14-15]和直角双曲线修正模型[16]进行了改进及修正,以便更好地分析光响应曲线,使其提取的相关光合参数更加准确,然而不同的拟合模型意义不尽相同,所以植物光响应曲线拟合模型的选择尤为重要,以避免产生较大的误差[17-22]。本文以金川荒漠植物疏花软紫草(Arnebiaszechenyi)、盐生草(Halogetonglomeratus)、木本猪毛菜(Salsolaarbuscula)为研究对象,比较其光合能力以及不同模型对同种植物和不同植物之间光响应曲线拟合结果,筛选出3种植物所对应的最优拟合模型,从而为当地植物光合作用对光的响应在植物生理和生态研究中的应用提供参考,更好地发挥其对当地水土保持、防风固沙等生态功能。
1 研究地区与方法
1.1 研究区概况
金川矿区位于甘肃中部,地处东经101°04′35″~102°43′40″,北纬37°47′10″~39°00′30″。以大陆性温带干旱气候为主,太阳辐射高,干旱少雨,昼夜温差大,蒸发量大,年均气温4.8 ℃,年均日照时数2878 h,无霜期141 d,多年平均降水量在110 mm 左右[23]。
1.2 实验方法
1.2.1 光响应测定
2017年8月中旬,采用Li-6400XT(Li-COR6400 Inc. USA)便携式光合仪在晴朗天气9:00—12:00进行实验。选择生长健康的疏花软紫草(Arnebiaszechenyi)、盐生草(Halogetonglomeratus)、木本猪毛菜(Salsolaarbuscula)各3株,选取中上部成熟叶片进行光响应测定[24]。设定光照强度为0、20、50、100、200、300、400、600、800、1000、1200、1400、1600、1800和2000 μmol/m2·s,流速为500 μmol/s,CO2浓度为400 μmol/mol。根据实测净光合速率随光强的变化情况,得到植物光响应曲线的表观量子效率、光补偿点、光饱和点等光合指标[25-26]。
1.2.2 光响应曲线拟合模型
1)直角双曲线模型[12]
(1)
2)非直角双曲线模型[13]
(2)
3) 指数模型[14-15]
(3)
4) 直角双曲线修正模型[16]
(4)
以上4个公式中:α为初始量子效率,Pnmax为最大净光合速率,Pn为净光合速率,I为光合有效辐射,Rd为暗呼吸速率,θ则表示光响应曲线弯曲程度的参数,取值范围为0<θ<1,β为光抑制项,为修正系数;γ为光饱和项,其值为α/Pnmax。
模型中的Pnmax、LCP、LSP等参数的计算见文献 [12-13]。
在模型(3)中,以0.99Pnmax所对应的光照强度为其饱和点,Pn=0时的光强为光补偿点[14-15]。
1.3 数据处理
采用Excel 2010软件对数据进行处理及作图。
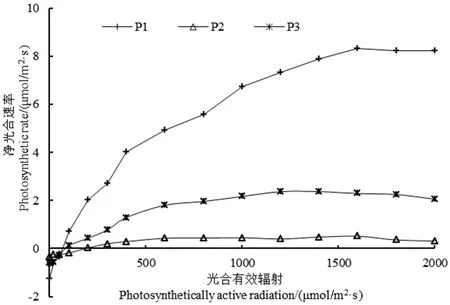
P1、P2、P3分别为疏花软紫草、盐生草、木本猪毛菜;下同
图13种植物的光响应曲线
Figure 1 Photosynthesis-light response curves of three plants
2 结果与分析
2.13种植物的光响应曲线及光响应模型拟合效果对比分析
由图1显示,3种植物的光响应曲线存在显著差异。Pn大小依次为疏花软紫草、木本猪毛菜和盐生草。当PAR小于200 μmol/m2·s时,随着光强的增加,3种植物的Pn均缓慢上升且差异较小,说明光照是限制光合的主要因素。在300 μmol/m2·s之后,随着PAR的增加至各植物达到光饱和点,其中,疏花软紫草、木本猪毛菜的Pn呈上升的趋势,分别在1600 μmol/m2·s、1400 μmol/m2·s时,Pn开始缓慢下降,出现了明显的光抑制现象;而盐生草随着PAR的增加,Pn趋于平稳,光抑制现象不明显,由此说明,3种植物对光照强度的需求大小依次为疏花软紫草、木本猪毛菜及盐生草。
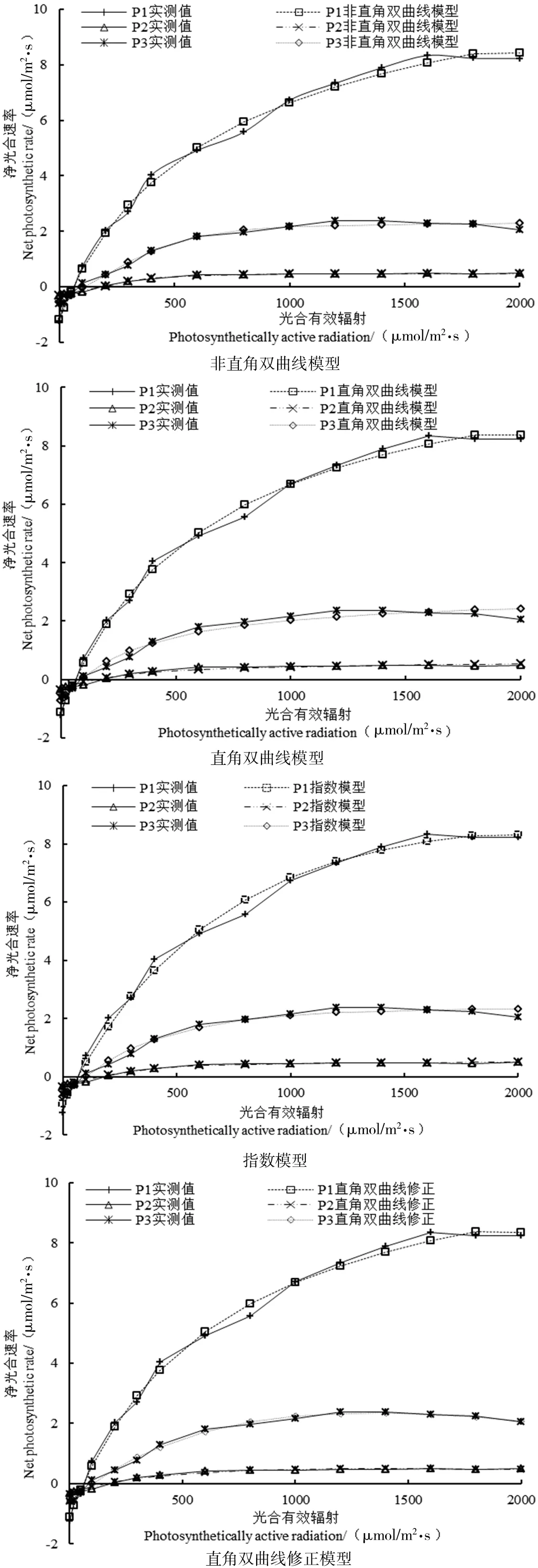
图2 4种模型对3种植物的光合-光响应曲线的模拟
由图2知,直角双曲线修正模型、非直角双曲线模型、指数模型对3种植物的拟合效果比较理想,R2均接近1。在PAR大于200 μmol/m2·s时,4种模型对3种植物拟合的光响应曲线变化基本一致,均呈逐渐增长的趋势,且这4种模型对植物光响应曲线的拟合值与实测值之间的差值相差均较小。当PAR大于400 μmol/m2·s时,疏花软紫草随着光照强度的增加,这4种模型之间的差异逐渐显著,其中非直角双曲线、指数模型、直角双曲线模型的Pn随着PAR的增加而升高,并逐渐趋向于平缓,而在直角双曲线修正模型的拟合下,Pn随着PAR的增加净光合速率有所下降。由此说明,疏花软紫草在直角双曲线修正模型的拟合下,出现光抑制现象,因此,对疏花软紫草拟合效果最好的模型是直角双曲线修正模型;木本猪毛菜随着PAR的增加,这4种模型之间的差异逐渐趋于显著,在光强达到1400 μmol/m2·s时,Pn呈现出下降的趋势,木本猪毛菜出现光抑制的情况,因此,直角双曲线修正模型对木本猪毛菜拟合效果较好;而对于盐生草来说,拟合效果最佳的为非直角双曲线模。
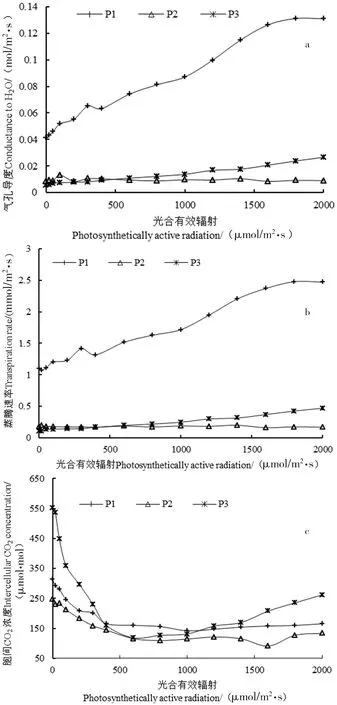
图3 3种植物的光合生理参数对比分析
2.2 3种植物的不同响应模型拟合参数对比分析
有研究表明:单纯的用决定系数来判断模型的适用性存在一定的缺陷,因此要评价某一模型对某种植物是否适用,不能仅仅依靠R2的大小来判断,更应该结合模型的最大净光合速率、光补偿点等参数与实际测量值之间的差异对模型进行综合判断[27-29]。由表1显示,疏花软紫草和木本猪毛菜在直角双曲线修正模型的拟合下决定系数最大,分别为0.9973、0.996;而盐生草在非直角双曲线模型的拟合下决定系数最大,为0.997。其中3种植物在4种模型的拟合下,其初始量子效率α始终均在(0≤α≤0.125)范围之内。就疏花软紫草而言,直角双曲线修正模型和指数模型与实测值的最大净光合速率最为接近,但指数模型的光饱和点远大于实测值,且决定系数R2为0.995,暗呼吸速率与光补偿点的拟合值和实际测量值相差不大。所以综合分析得知,疏花软紫草的最优拟合模型为直角双曲线修正模型。盐生草的最大净光合速率与实测值之间最为接近的模型为非直角双曲线模型和指数模型,这两种模型对盐生草的光补偿点的拟合值和实测值差异不显著,但指数模型的光饱和点远小于实测值。且决定系数为0.990,由此可知,非直角双曲线模型为盐生草的最优模型,对盐生草的拟合度最好。对木本猪毛菜而言,直角双曲线修正模型和指数模型拟合的光合参数较为接近,但指数模型的光饱和点和暗呼吸速率远大于实际测量值,且指数模型的决定系数R2为0.989,而直角双曲线修正模型的决定系数R2为0.996。决定系数R2越靠近1,说明该模型对该种植物的拟合程度越高,但并不能说明就与实测值完全相符[30],即出现过拟合的现象,其原因可能是光响应测量时设置的光强梯度太大造成的,因此直角双曲线修正模型拟合效果要优于指数模型。
2.3 不同植物光合相关参数对比分析
绿色植物净光合速率、蒸腾速率、胞间CO2浓度、气孔导度等光合生理参数及环境因子如大气温度、空气相对湿度、光照条件等紧密相关。由图3-a、b显示:疏花软紫草的蒸腾速率与气孔导度随着光合有效辐射的增加呈先上升后逐渐趋于平缓;木本猪毛菜的气孔导度与蒸腾速率随着光合有效辐射的增加呈缓慢上升的趋势;而盐生草随有效光辐射的增加气孔导度与蒸腾速率、胞间CO2浓度、净光合速率整体呈缓慢下降的趋势,这可能与该地区重金属含量有关[31-32]。由图3-c显示:3种植物的胞间CO2浓度随光照条件的增加变化趋势基本一致,均呈先降低后缓慢上升的趋势,且木本猪毛菜变化幅度较大,而疏花软紫草和盐生草的变化相对比较平缓。
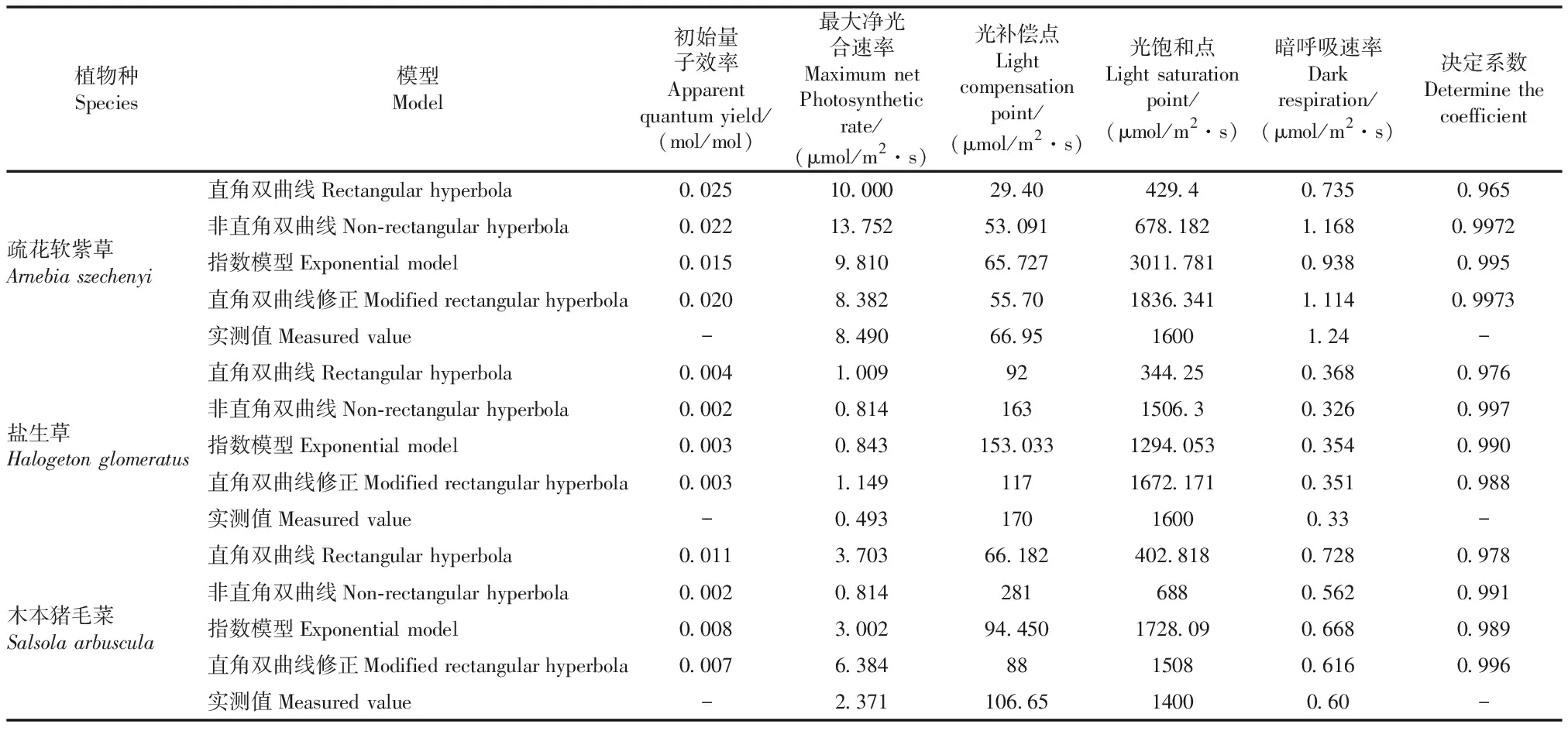
表1 4种模型拟合光响应曲线参数
注:-表示不存在
3 讨论
通过测定3种植物的光响应来获取Pnmax、LSP和LCP 3个重要指标,发现3种植物的光合能力及对光强的适应存在着明显的差异,其光合能力大小依次为疏花软紫草、木本猪毛菜和盐生草,其原因可能与植物自身的形态结构特征等生理功能有关[33]。本文采用4种不同光响应模型对3种植物光动态响应进行拟合,显示其拟合曲线和光合参数均存在显著差异。
非直角模型、指数模型、直角模型、直角双曲线修正模型是常见的光响应拟合模型。已有报道非直角模型、直角模型拟合得到的LSP偏低、Pnmax偏高,不能解释光饱和区域的光抑制现象,指数模型拟合得到的Rd偏低等已得到大家的公认[34-35]。有研究指出,直角双曲线修正模型拟合的各项参数较好[36],这与本研究中疏花软紫草和木本猪毛菜的研究结果一致,直角双曲线修正模型拟合的曲线符合植物生理生长规律,并且R2及拟合参数要比其他3种模型拟合较好,精确度较高,拟合效果最优。而非直角双曲线模型对盐生草的拟合效果最佳,这与段爱国等[37]的研究结果相一致,他指出直角双曲线修正模型仅适用于光抑制现象的光响应曲线的拟合,至于直角双曲线修正模型为什么不适用于拟合无光抑制现象的光响应曲线,还有待于进一步研究。
这4种模型对同一种植物的光响应曲线拟合结果存在较大差异,同一种模型对不同物种的拟合亦存在显著差异。分析植物的光响应时,单纯的用决定系数来判断模型的适用性存在一定的缺陷,这与叶子飘等[9]的研究结果一致,应考虑所选模型对该种植物的光响应特征是否适合,从几个模型中选出拟合效果最优的一个或综合几个模型的结果加以应用,这样才能达到最佳的效果,并提高对模型中各参数估计的准确性。
4 结论
1) 通过对金川矿区3种植物光合参数的测定和拟合分析表明:直角双曲线修正模型对疏花软紫草和木本猪毛菜的拟合效果最好,较好地反映了植物受光抑制的现象,而非直角双曲线模型对盐生草拟合度最好。
2)3种植物光合能力存在明显的差异,4种模型对同一植物的光响应曲线拟合结果存在较大差异,同一模型对不同物种的拟合亦存在明显差异。在光合有效辐射较低时,3种植物的净光合速率相差较小,随着有效光辐射的增加,其净光合速率的差异逐渐显著,光合能力依次为:疏花软紫草>木本猪毛菜>盐生草。
3)疏花软紫草适合在光照强度相对比较高的环境中生存,木本猪毛菜次之,而盐生草则对光照条件要求相对较低,适合在光照强度比较低的环境中生存。
