红蓝绿LED延时补光对日光温室番茄育苗的影响
2019-10-16朱鹿坤陈俊琴赵雪雅张传玲王景瑞张军超齐明芳
朱鹿坤 陈俊琴 赵雪雅 张传玲 王 冲 刘 曼 王景瑞 张军超 齐明芳
(沈阳农业大学园艺学院,北方园艺设施设计与应用技术国家地方联合工程研究中心,设施园艺省部共建教育部重点实验室,辽宁沈阳 110866)
我国北方冬春季节光照强度较弱,日光温室采取的防寒保温措施又不同程度地缩短了光照时数,光照问题成为日光温室果菜类蔬菜秧苗生长发育不良和质量下降的最突出因素。同时,日光温室集约化育苗普遍存在光照强度低而温度相对较高的现象,易导致植株营养生长过旺,从而造成蔬菜幼苗生长不良和徒长的问题(李天来,2013)。如何在育苗过程中进行合理补光,成为生产中亟待解决的问题。
光为植物光合作用提供必要的能量,还可作为信号在植物光形态建成方面起到调节作用(孙洪助 等,2015)。植物光合作用所需光的波长在400~700 nm之间,其中红光与蓝光部分对植物的生长发育调控作用最显著(李倩,2010)。红光具有抑制节间伸长、增加分蘖、促进光合色素的形成、增强光合作用并增加糖分积累、提高抗逆性等作用(Wang et al.,2010;许大全 等,2015;周成波 等,2017);蓝光具有抑制茎伸长、促进叶绿体发育并提高叶绿素含量、增加叶片厚度、调节气孔开放、促进根系发育、调控植株的向光性等作用(陈祥伟等,2013);而绿光部分也能被植物叶片吸收利用,能够提高植株光合速率和干物质积累,其中短波绿光更有益于植株生长(何蔚 等,2016)。绿光由于高透射性能够照射到植株下层,可刺激较低冠层利用光能,同时还能延缓叶片衰老,进而促进光合作用(Materová et al.,2017)。Bouly等(2007)发现绿光能够参与植物光合作用,影响叶片气孔的开放,但不会降低植物的光合能力,对光合作用具有一定积极的作用。绿光还能提高叶用莴苣(生菜)的干质量、叶面积和可溶性糖含量,保持叶片厚度,增加植株叶绿素含量,提高吸收光的能力,改善生菜的品质(王晓晶 等,2019)。
LED(light emitting diode)用于作物补光已成为研究的热点,LED光质对植物生长发育和光形态建成具有很大的潜力。研究表明,对设施蔬菜生产中的嫁接育苗进行补光,可促进胚轴伸长以达到嫁接水平,进而提高嫁接苗的成活率,移栽后的嫁接苗补照红蓝组合光,能促进嫁接伤口的快速愈合,提高对环境的适应能力(Jang et al.,2013)。黄瓜、辣椒和番茄幼苗通过补照红蓝光,壮苗指数得到显著提高(Hernández,2016)。红蓝光可以促进叶用莴苣(生菜)生长,提高花青素含量(Lee et al.,2010)。不同光质进行组合对植物生长的促进作用优于单质光。赵飞和高志奎(2011)研究表明,在红蓝组合光的基础上补充绿光会显著提高黄瓜叶片中叶绿素的含量,增强光合能力。在红蓝组合光中添加绿光,绿光低于24%时能促进小麦生长,但含量过高会抑制生长(Goins et al.,1997)。
利用LED光调控技术培育健壮的蔬菜幼苗是一项有效的方法,但目前对LED光谱选择与组配优化方面的研究仍显不足。近年来,补光一般以红蓝组合光为主,对绿光的研究较少。本试验利用LED红蓝绿组合光对番茄幼苗进行夜间延时补光,探讨补光对番茄幼苗的调控作用,筛选促进番茄幼苗健壮生长的最佳组合光,以期为LED光源在番茄设施育苗的应用与理论研究上提供实践指导。
1 材料与方法
1.1 供试材料
采用广东威谱照明科技有限公司生产的LED灯作为补照光源,按试验要求制成LED植物生长灯(40 cm×30 cm×7 cm),额定电压为220 V,额定功率为15 W,补光的光照范围为高25 cm、宽50 cm、长80 cm之间。光谱的测定采用地物光谱仪GER1500型(Ocean Optical,美国),光照强度采用Li-250型光强仪(LI-COR,美国)进行测定。各光质峰值波长见图1,LED红光(R)峰值波长为630 nm,LED蓝光(B)峰值波长为460 nm,LED绿光(G)峰值波长为520 nm。
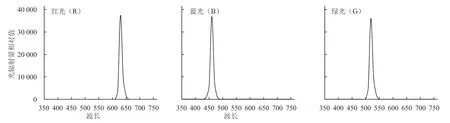
图1 LED红、蓝、绿光谱图
1.2 试验设计
试验于2017年10月15日至2018年5月26日在沈阳农业大学园艺学院蔬菜科研基地8号日光温室内进行,共3次重复。供试番茄(Solanum lycopersicumL.)品种为辽宁省农业科学院提供的辽园多丽。种子催芽处理后,播于50孔穴盘中进行育苗,待幼苗第1片真叶露出时进行补光处理。LED组合光采用白光(W)以及由红光(R)、蓝光(B)、绿光(G)3种LED单质光进行6种不同比例搭配的组合光。灯板设计主要以红灯为主,中间按一定比例均匀分布着蓝灯及绿灯,其中各组合比例(表1)分别为R9B1(9∶1,灯的数量比,下同)、R8B2、R7B3、R9G1、R8B1G1、R7B2G1。各LED组合光光谱成分见图2。调节光源与幼苗的距离使光照强度达到60 μmol·m-2·s-1。每种灯源相距1 m排列放置,各处理之间装置黑白反光膜,以防光源之间相互干扰。所有幼苗白天放置于日光温室中正常生长,夜间进行延时补光,以不补光处理作为对照。幼苗的培养条件为白天温度23~28 ℃,夜间12~18 ℃,补光处理延时补光时间为4 h·d-1,补光时段为17:00~21:00,每处理50株,2次重复。待补光处理30 d后取样测定相关指标。
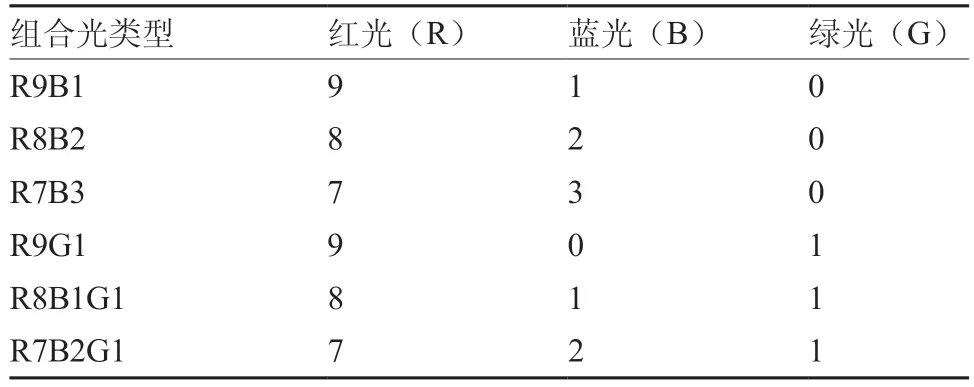
表1 不同比例LED组合光类型
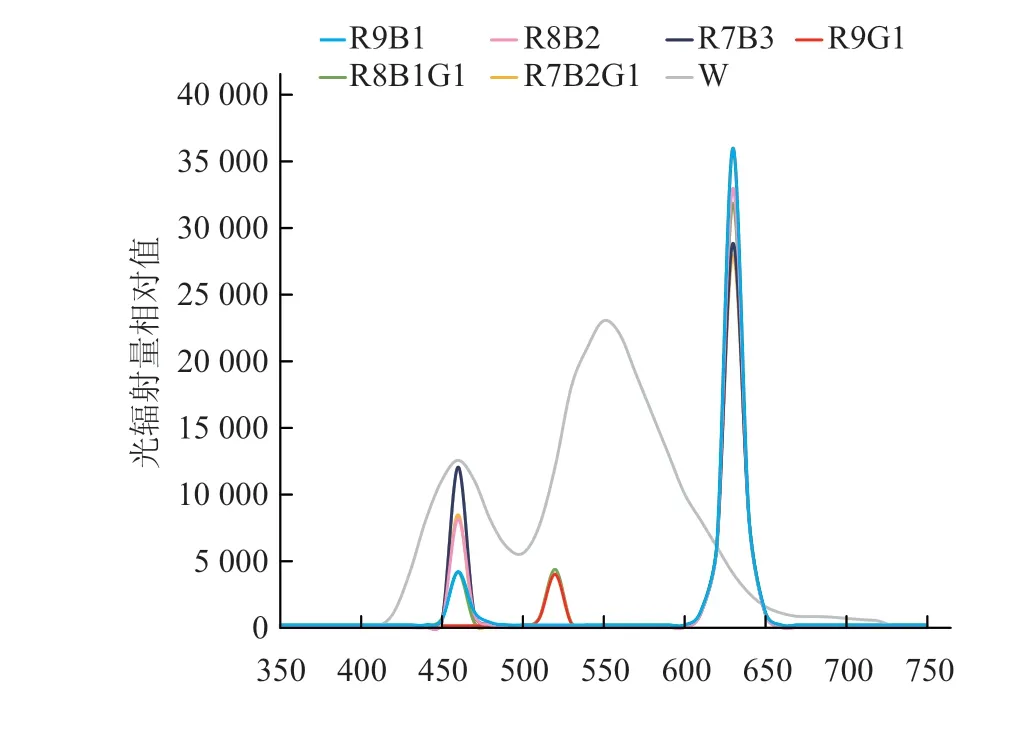
图2 LED组合光光谱图
1.3 测定项目及方法
株高使用直尺测量幼苗从地上根基处至生长点的距离;茎粗用游标卡尺测量幼苗的子叶基部下1 cm处的部位;叶面积采用叶片长×宽×叶片形状系数的计算公式,叶片形状系数采用称重法测定(侯福林和范海,2015),本试验测得番茄叶片形状系数为0.541;将幼苗根部用水清洗干净后,吸干表面水分,用电子天平测定植株地上部、根部鲜质量,然后进行烘干处理,称取干质量;并计算壮苗指数、根分配率、G值、叶面积/株高和比叶面积。

采用氯化三苯基四氮唑TTC法测定幼苗的根系活力;采用丙酮和乙醇1∶1(体积比)浸泡法提取叶绿素,用紫外光分光光度计U-5100测定叶绿素a、叶绿素b含量,并计算叶绿素a+b和a/b值。采用LI-6400型便携式光合测定仪(LI-COR,美国),测定幼苗的相关光合参数,包括净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci);采用蒽酮比色法测定幼苗叶片中可溶性糖含量。
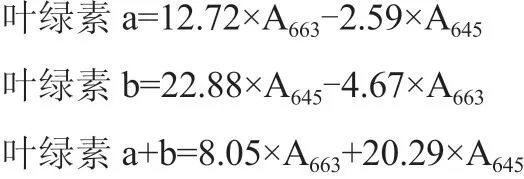
1.4 数据处理
采用Excel 2010软件进行数据处理和作图,采用SPSS 22.0软件对3个重复的试验数据平均值进行统计,采用Duncan检验法进行多重比较和差异显著性分析。图表中的数据为平均值±标准差。
2 结果与分析
2.1 不同组合光对番茄幼苗形态指标的影响
番茄幼苗进行补照LED组合光(表2、图3),白光(W)和组合光处理均有提高幼苗茎粗、叶面积的趋势。其中,组合光R8B1G1和R7B2G1处理对番茄幼苗茎粗增长作用最显著,相比对照分别提高了23.50%、27.42%。在株高方面,除R9G1处理外,其他组合光及白光均抑制了番茄的株高,高成分蓝光的抑制效果尤为明显。多数补光处理有利于提高幼苗鲜、干质量,其中R7B2G1组合光对全株鲜质量和干质量的影响最显著,比对照分别提高了17.84%和30.91%。以上结果表明,补光在一定程度上促进了番茄幼苗生长,不同比例的光质处理差异显著,其中R7B2G1处理总体效果较好。
2.2 不同组合光对番茄幼苗壮苗指标的影响
从表3可以看出,番茄幼苗补照LED组合光可有效提高幼苗的壮苗指标,从而达到壮苗的效果。在壮苗指数方面,白光和红蓝绿组合光处理的番茄幼苗壮苗指数均得到了提高,其中组合光R8B1G1和R7B2G1处理比对照分别显著提高了36.96%、57.61%。同时,R7B2G1处理显著提高了幼苗的G值、叶面积/株高、根分配率以及比叶面积,说明红蓝绿组合光特别是R7B2G1处理较适合番茄幼苗的生长,可以达到壮苗的效果。番茄幼苗补照LED组合光能提高其根系活力。在红蓝组合光处理中,随着蓝光成分的增加番茄幼苗的根系活力也随之增加;在红蓝组合光中加入绿光成分,番茄幼苗的根系活力得到提高,其中R7B2G1处理比对照显著提高了48.32%,但R9G1处理低于红蓝组合光处理,说明绿光对番茄幼苗根系活力的促进作用需要红蓝光作为基础。
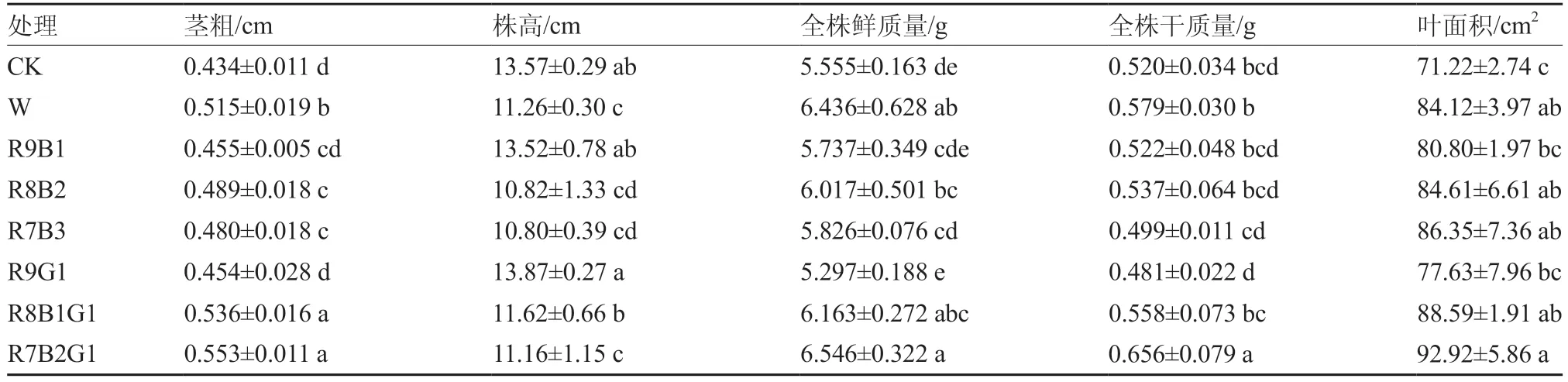
表2 不同LED组合光下番茄幼苗的生长形态

图3 番茄幼苗延时补照LED组合光的生长形态
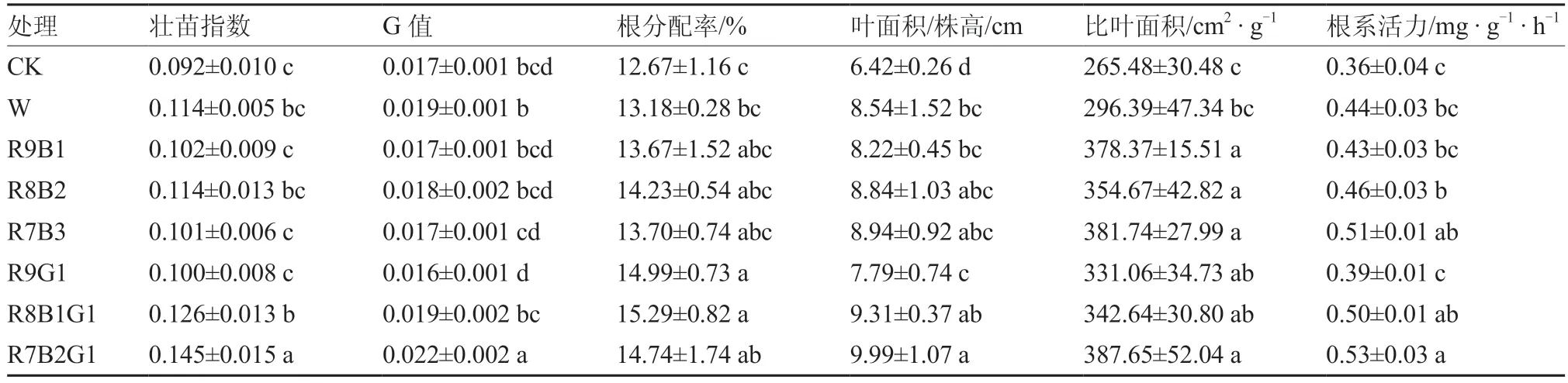
表3 不同LED组合光下番茄幼苗的壮苗指标
2.3 不同组合光对番茄幼苗叶绿素含量和光合速率的影响

图4 不同LED组合光下番茄幼苗的叶绿素含量
从图4可看出,番茄幼苗补照LED组合光,叶绿素a含量随着蓝光比例的增加而增加,特别是组合光R7B3和R7B2G1处理含量较高,由此可见,红蓝光特别是其中加入绿光时,有利于促进番茄幼苗叶绿素a含量的增加。各补光处理番茄幼苗的叶绿素a、叶绿素b和叶绿素a+b含量均显著高于对照。W处理番茄幼苗的叶绿素a和叶绿素b含量与对照差异不显著。在叶绿素a/b方面,除W和R9G1处理外,其余补光处理均显著低于对照。综上可知,对番茄幼苗补照LED组合光能够提高叶绿素的含量,促进植株吸收光能。
从表4可以看出,在净光合速率和蒸腾速率方面,各LED补光处理均显著高于对照,其中R7B2G1处理的净光合速率和蒸腾速率最高。在番茄幼苗气孔导度方面,除W处理与对照差异不显著外,其余补光处理的气孔导度均显著高于对照,其中R7B2G1处理气孔导度最大。在组合光处理中,随着蓝光比例的增加,气孔导度也在增加,说明蓝光对番茄幼苗气孔的开放具有促进作用。在胞间CO2浓度方面,各补光处理均低于对照,而且随着蓝光比值的升高,胞间CO2浓度持续下降。由此可见,红蓝绿组合光在一定程度上具有促进番茄幼苗利用叶片中CO2的作用,从而提高植株的光合能力。
2.4 不同组合光对番茄幼苗可溶性糖含量的影响
从图5可以看出,除了R9G1处理外,各补光处理叶片中可溶性糖含量均显著高于对照,其中R8B1G1和R7B2G1处理的可溶性糖含量分别比对照显著提高64.52%和45.51%。在组合光处理中,随着蓝光比例的增加,可溶性糖含量随之减少。

表4 不同LED组合光下番茄幼苗的光合指标
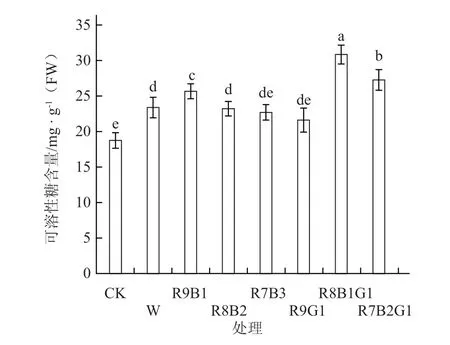
图5 不同LED组合光质下番茄幼苗叶片的可溶性糖含量
3 结论与讨论
光质在一定程度上调节着植株的生长和发育,红蓝光能显著促进黄瓜、辣椒、番茄的生长(崔瑾 等,2008)。本试验结果表明,红蓝绿组合光对番茄幼苗的茎粗、全株鲜干质量与叶面积有明显的促进作用,与前人研究结果一致。红蓝组合光在一定程度上抑制了幼苗的株高,红蓝绿组合光处理中番茄幼苗的株高也明显降低,与令狐伟等(2015)研究发现采用绿光补光能抑制弱光条件下幼苗徒长的结果有相似之处。本试验中,红绿组合光的番茄幼苗株高显著高于红蓝绿组合的株高,表明蓝光在控制株高方面有积极的作用。番茄幼苗补照红蓝绿组合光能显著提高壮苗指数,组合光R7B2G1还显著提高了番茄幼苗的G值,叶面积/株高和比叶面积,主要原因可能是红蓝绿组合光改善了植株的整体长势,特别是增加了茎粗、叶面积和干物质的积累,进而促进了幼苗的健壮生长。另外,在根系活力方面,随着蓝光成分的增加,番茄幼苗的根系活力存在上升的趋势,这与蓝光能提高茄子和辣椒幼苗根系活力的研究结论一致(胡阳 等,2009);红蓝绿组合光R7B2G1处理的幼苗根系活力最大,说明该组合在促进番茄幼苗生长的同时也有效地改善了地下根系的生长状态,增强了根系吸收水分和营养物质的能力;也有可能是因为红蓝绿组合光的共同作用促进了同化物质向地下部的分配而改善了根系的生长。可见,在红蓝光组合中适当增加一定比例的绿光成分能进一步促进番茄幼苗的健壮生长。
光合色素是植物光合作用的物质基础,其含量与组成直接影响着光合作用(郑洁 等,2008)。本试验中,红蓝绿组合光显著提高了叶绿素a、叶绿素b以及叶绿素总含量,可以反映出红蓝绿组合光的互补作用在促进叶绿素合成和提高其含量方面较为明显,与赵飞和高志奎(2011)在黄瓜上的研究结果相似,说明番茄幼苗在红蓝绿组合光处理下含有较高的叶绿素含量可能是提高光合速率的重要因素。
本试验中,R7B2G1处理的净光合速率、蒸腾速率、气孔导度均显著高于对照,同时该处理的壮苗指数、比叶面积和全株鲜、干质量也最大,表明适当增补绿光对促进番茄光合作用和提高光合产物的积累也有显著作用。红蓝光可以通过提高光合速率来促进植物的生长发育,其原因是红光与蓝光的光谱与叶绿素吸收光谱较为一致(王丽伟 等,2017)。而红蓝组合光中加入绿光后幼苗的光合作用有了进一步提高,其原因可能是红蓝绿组合光与红蓝组合光相比能提高幼苗的光合酶活性和光化学效率,改善光系统Ⅱ性能,进而提高净光合速率和光合电子传递速率(文莲莲 等,2018)。在组合光处理中,随着蓝光比例的增加,气孔导度也在增加,表明蓝光对番茄幼苗气孔的开放具有促进作用(蒲高斌 等,2005)。各补光处理的胞间CO2浓度均低于对照,而且随着组合光中蓝光比例的升高,胞间CO2浓度在持续下降,在红蓝组合光中加入绿光后,胞间CO2浓度降幅更大,表明蓝光和绿光能有效促进叶片中CO2在光合作用下的利用和转化,来增加干物质的积累。
本试验的红蓝组合光处理中,随着蓝光比例的增加,番茄叶片可溶性糖含量随之减少,与王丽伟等(2017)蓝光可以降低可溶性糖含量的研究结果一致。在红蓝绿组合光处理中,番茄幼苗叶片中可溶性糖含量比红蓝组合光处理显著增加,表明绿光对增加叶片中可溶性糖含量具有促进作用,分析其原因可能是绿光能够缓解高红光成分造成的光抑制,进而增加叶片碳水化合物的合成与积累。
综上所述,对番茄幼苗进行延时补照红蓝绿组合光,通过提高幼苗叶片的叶绿素含量、净光合速率以及根系活力,进而增加茎粗、叶面积和全株鲜、干质量,可以有效改善其生长形态,达到壮苗的效果,其中R7B2G1处理效果最佳。本试验为现代工厂化育苗生产提供了理论依据,关于红蓝绿LED光质对番茄幼苗的具体作用机理及途径仍待继续深入研究。
