不同叶背补光模式对戈壁温室番茄叶片光合性能与固碳效应的影响
2019-10-16蒋程瑶李玉姗
蒋程瑶 宋 羽 李玉姗
(1西北农林科技大学园艺学院,陕西杨凌 712100;2新疆农业科学院农作物品种资源研究所,新疆乌鲁木齐 830091)
光为植物生命活动提供能量,同时也是影响植物生长发育的重要环境因子。提升作物光环境,不仅能提升作物生物量积累,更能影响产量和品质(邵丽 等,2018)。因此,针对不同作物的光环境需求的探索,一直是设施园艺领域的研究热点。新疆南部多戈壁、沙漠,农业用地少。近年来,在国家和自治区政府的大力支持下,南疆四地州设施产业发展迅速,根据新疆维吾尔自治区农业农村厅的统计数据,截至2017年12月,该地区设施农业总面积约占全疆设施农业总面积的1/3,约2.09万hm2(邹平 等,2019)。然而南疆地区冬春和晚秋季节光照时间短,特别是春季多沙尘和连阴天,设施温室内严重缺光,作物正常生长严重受限(宋羽,2017)。利用人工光源在温室内对作物进行补光,是保证植物正常生长,促进高产保质,提高设施经济效益的有效途径。目前,已有大量研究从光源选择(杨其长,2008;Lu et al.,2012)、光质(Lu et al.,2012;Song et al.,2016;Jiang et al.,2018)、光强(王君 等,2016;宋羽,2017)、光周期(薛国萍 等,2017)等多个角度进行设施补光研究,关于多个光环境因子共同作用的研究也见诸报道(程亚娇 等,2018;何蔚 等,2018)。
番茄(Solanum lycopersicumL.)是南疆重要的设施蔬菜,已有研究表明叶背补光能够显著提高番茄叶片光合效能,提升果实产量与品质,获得更高的经济效益(Song et al.,2016;Jiang et al.,2018),但对更经济的补光策略选择仍然缺乏系统研究。本试验以目前市售植物用LED灯管为光源,以戈壁温室生产番茄为供试材料,研究不同光强和补光时间耦合的叶背补光处理下,番茄生长、光合特性及固碳效益的响应,以期为南疆设施蔬菜的高产优质生产提供技术支持。
1 材料与方法
1.1 试验材料
以广州南蔬农业科技有限公司的番茄品种NS3389为试验材料;采用商品基质栽培(培蕾2号,培蕾有机肥料有限公司);试验使用LED灯(宸华照明有限公司),光源板由635 nm波长的红色(R)LED灯珠、480 nm波长的蓝色(B)LED灯珠,以及全波长白色(W)LED灯珠按照1∶2∶3的比例组合而成,其光谱分布特性如图1-a所示,不同波谱均在距离灯正下方20 cm处测定。
1.2 试验处理

图1 试验所用补光灯光谱分布(a)与温室内补光现场(b)
试验于2019年1~7月在南疆阿克陶日光温室产业园中进行。番茄采用基质袋培,行距0.8 m,株距0.2 m,定植密度为6.25株·m-2。根据宋羽(2017)的试验方法,LED补光灯设置在底部冠层距叶片10 cm处(第1穗果摘叶,果下保留3片叶),光处理集中在坐果后—转色开始时期。补光周期设定为当地时间(下同)6:00~22:00(14 h),以无补光处理为空白对照(CK),并设置T1处理:14 h持续低光强补光(补光光强 为100 μmol·m-2·s-1);T2处 理:揭 帘(平 均8:00)前与盖帘(平均17:30)后高光强补光(补光光强为200 μmol·m-2·s-1),补光灯的开关由卷帘机的运转决定,补光时间大致为6:00~8:00和17:30~22:00;T3处理:补光周期内揭帘后室内光强低于150 μmol·m-2·s-1时自动补光(补光光强为100 μmol·m-2·s-1)。补光装置由时间和保温被电机共同控制模块进行自动控制,并自动记录开灯时长,各处理单独设电表进行用电统计,光处理现场如图1-b所示。
单个试验区内,植株密度为12株·m-2,每个处理包括1排、20株植株,各处理间隔1排、20株植株,避免组间干扰,边界设62株植株防止边界效应,共设3个重复试验区。试验期间,温度变化范围为25~30 ℃(昼)/10~15 ℃(夜),湿度变化范围为70%~90%,CO2变化基本与外界一致。
1.3 指标测定
1.3.1 光合参数及叶绿素荧光参数测定 参照宋羽(2017)的方法,在补光处理期间每隔7 d对底部冠层代表性叶片采用Li-6400XT便携式光合 系 统(Li-Cor Inc.,Lincoln,NE,USA),于9:00~14:00期间测定一次叶片光合参数,共进行15次测定。每次测定每个处理选择3株植株,每株选取同样高度的3个采样点。测定参数包括净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)。同时分别测定各处理冠层光响应曲线,并计算表观叶肉细胞导度(Gm)(Fischer et al.,1998)。测定叶绿素荧光参数,分别计算暗适应下PSII最大量子产率(Fv/Fm),光适应下的PSII最大光化学效率(),PSII电子传输的量子产率(ФPSII),及非光化学淬灭(NPQ)。
1.3.2 番茄果实变化与产量 记录植株开花时间,拍照记录果实颜色变化,每个处理随机选取第2穗果20个成熟度一致的果实,称量其单果质量(刘露,2018)。共留5穗果,最后统计所有果实鲜质量,计为产量。
1.3.3 植株固碳效益比较 参照宋羽(2017)的方法,对光处理14 d后的植株进行13C饲喂处理3 h,每个处理随机选3株植株,取样3次重复,由稳定同位素质谱仪测定功能叶(果实周围3片叶)、果实内13C含量(Integra 2,SerCon Ltd,UK)。
1.3.4 植株生长形态 果实收获后,测量番茄植株株高、茎粗,测定叶面积、地上部(茎、叶)干鲜质量、根干鲜质量等指标(宋羽,2017),计算植株健康指数(Fan et al.,2013)和比叶重(Hernández &Kubota,2016)。

1.3.5 经济效益分析 参照宋羽(2017)的方法,计算电能效率和投入回报率,分析各补光处理的经济效益。

1.4 数据处理
试验数据采用SPSS 11.0软件(SPSS Inc.,Chicago,IL,USA)进行统计分析。
2 结果与分析
2.1 不同补光处理番茄叶片光合参数分析
从表1可以看出,补光处理下的番茄叶片净光合速率(Pn)较对照显著提高,其中T1处理较对照提升52.6%,T2和T3处理较对照提升约40%。气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)和表观叶肉细胞导度(Gm)的变化趋势相近,T1、T2和T3处理下的各参数值无显著差异,均显著高于对照。通过绘制光响应曲线,可见在测试时间内,Pn-PPFD变化趋势与Pn相同(图2):T1处理下叶片达到饱和光强下的最大Pn最高,较对照提升61.6%;T2和T3处理次之,提升幅度接近46.7%。

表1 不同补光处理对番茄叶片光合参数的影响
2.2 不同补光处理番茄叶片叶绿素荧光特性分析
由表2可知,各补光处理番茄叶片Fv/Fm无显著差异。T1处理叶片的Fv′/Fm′显著高于对照和T3处理,T1和T2处理叶片的ФPSII和NPQ较大,显著高于对照,但两者之间无显著性差异。
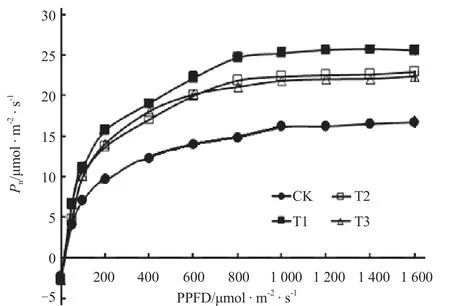
图2 不同补光处理下番茄叶片的光响应曲线

表2 不同补光处理对番茄叶片叶绿素荧光特性的影响
2.3 不同补光处理番茄果实变化与产量分析
从表3和图3可以看出,补光处理的番茄果实成熟时间明显提前,定植后32 d,补光处理的植株就开始开花,定植后81 d,T1处理的果实开始变为红色,其他处理的果实仍处于青熟期,T2处理果实的成熟时间仅次于T1处理。定植后84 d,T1、T2处理的果实已基本完成转色,T3处理开始转色,而对照果实仍处于青熟期,定植后94 d对照才完全成熟。各补光处理的番茄单果质量和产量明显提高,其中T1和T2处理的单果质量和产量均显著高于对照。
2.4 不同补光处理番茄叶片固碳效应分析
分别以果实周围3片叶为13C碳源,测得48 h后植株各部位的13C含量分数(图4)。总体上讲,补光处理的番茄植株叶片光合产物在果实内积累量比重高于对照。果上和果下第1片叶作为13C碳源时,光合产物在果实内积累量较高。对照果下第3片叶光合产物基本滞留在本叶,输出用于果实干物质积累的部分占比仅22.1%。T1和T2处理的叶片光合产物用于果实干物质形成的部分在各叶片中占比均较高,且远离果穗的衰减幅度较小,固碳效果稳定性好。

表3 不同补光处理对番茄植株开花时间和果实产量的影响

图3 不同补光处理下番茄果实转色情况

图4 不同叶片作为碳源时不同补光处理下植株13C光合产物的输出分布
2.5 不同补光处理番茄植株形态指标分析
从表4可以看出,与对照相比,各补光处理的番茄地上部干质量、根干质量、植株健康指数、比叶重均显著高于对照,株高和茎粗各处理与对照差异均不显著。除植株健康指数和比叶重外,3个补光处理间各指标无显著差异。
2.6 不同补光处理经济效益分析
对各补光处理的耗电量进行统计(表5),T1处理的耗电量显著高于T2和T3处理,产量增长量也表现出相同的趋势。但是代入番茄批发价格和补光灯投入成本后,T1和T2处理的电能效率和投入回报率差异不显著,但均显著高于T3处理。说明T1和T2这两种补光方式能获得较高的经济效益。

表4 不同补光处理对番茄植株生长的影响

表5 不同补光处理的能源效率
3 结论与讨论
光环境直接影响植物生长发育,由于沙尘、雨雪以及连阴等不利天气条件造成的设施内弱光寡照,不仅会抑制植物生长,限制作物生产潜力,更有可能会导致病害发生,威胁产品安全,进而导致设施的经济效益降低(邵丽 等,2018)。通常番茄中、低部冠层的光量非常低,而直接对这部分冠层进行补光,且补光光强在100~200 μmol·m-2·s-1时能够较好地促进番茄叶片光合参数的提升且避免补光过剩(宋羽,2017)。基于此研究结果,并结合LED灯具成本,本试验设置了100 μmol·m-2·s-1持续补光(T1、T3)和200 μmol·m-2·s-1间歇补光(T2)的补光模式。试验结果表明,持续每天100 μmol·m-2·s-1的补光处理下(T1)的叶片瞬时Pn显著高于其他处理,同时光响应曲线也表明T1处理下叶片对光强的响应速度快,饱合光强下的最大Pn也显著高于其他补光处理,说明这种补光模式下叶片的光合潜能最为优良。另一方面,结合Gs、Ci和Gm的变化趋势一致,且与Pn变化相近这一结果,说明戈壁温室内番茄光合作用的限制因素主要是气孔因素(Deans et al.,2019),即叶片气孔的张开程度决定了CO2的进入,供给光合原料,而非提升CO2的羧化效率来促进Pn的提升。这与前人关于叶背补光提升番茄光合效能的作用方式的研究结果并不完全一致(Song et al.,2016),出现这一差异,推测是由于试验条件差异(室内湿度差异)造成。同时这也说明,在戈壁温室番茄生产期间,在补光的同时适时补充CO2将能够更有效地促进植株对光能利用。叶绿素荧光参数变化说明了植株在受胁迫状态下的应激反应,通常用作胁迫或修复的判定指标。其中Fv′/Fm′的降低意味着胁迫的存在,ФPSII表示光合作用的电子供应能力,直接影响Pn,而NPQ则可以表征叶片的光保护能力。本试验结果表明,T1与T2处理的叶片自身光保护能力要优于其他处理。但是T2处理中Pn显著低于T1处理,但ФPSII和NPQ与T1处理差异不显著,说明T2处理CO2的羧化效率与T1处理差异不显著,这也再一次印证了本试验中Pn的提升是由于气孔因素的作用。
叶片光合能效的提升使产品干物质积累能更有效地进行。基于前人对补光功能叶的判定(宋羽,2017),本试验中选择果周6片叶为研究对象,对不同补光处理的功能叶片固碳分配模式进行测定,表明补光处理能够显著提升功能叶的固碳水平,并维持较高的向果实转运的比重,与前人的研究结果基本一致(宋羽,2017)。T1和T2处理的叶片光合产物用于果实干物质形成的部分在各叶片中占比均较高,且趋势较为稳定,这也导致植株发育较好,开花快,果实转色时间提前,最终果实产量高。
在对生产措施的可行性评估中,成本是一个重要的参考指标。补光消耗的能源成本可能会阻碍这一措施的经济可行性。因此,现代温室作物生产系统需要节能高效的照明措施。通过对番茄产量、售卖价格、灯具成本等多因素进行计算,得出T1和T2处理的投入回报率基本持平,显著高于T3处理,说明在南疆设施番茄生产中既可以选择较低光强的持续补光模式,也可以选择揭帘前和盖帘后较高光强的延长光周期的补光模式,都具有较高的性价比。
