黑米亲本总黄酮与花色苷配合力及遗传力分析
2019-10-08程祖锌黄昕颖肖长春林荔辉郑金贵
程祖锌,黄昕颖,肖长春,2,林荔辉,2,许 明,郑金贵*
(1.福建农林大学农产品品质研究所,福建 福州 350002;2.福建农林大学作物科学学院作物遗传育种与综合利用教育部重点实验室,福建 福州 350002)
【研究意义】与普通稻米相比,黑米具有较高的营养和保健价值[1-3],黑米的核心成分为花色苷——以黄酮核为基础的一类物质,具有抗氧化、抗衰老、降血脂、抗炎以及抑制肿瘤生成等功能[4-6]。但不同黑米品种之间总黄酮、花色苷含量差异极显著,含量高低不一[7]。选育高黄酮、高花色苷的黑米品种对提高黑米的营养和保健价值具有重要意义,杂种优势利用是提高黑米功能成分性状的有效途径,配合力法是杂种优势的主要测量方法。因此,研究黑米两系杂交稻亲本总黄酮、花色苷含量的配合力及遗传力,对合理组配高总黄酮、高花色苷黑米两系杂交稻组合具有重要意义。【前人研究进展】育种工作者越来越重视特种稻黑米良种的选育,已育成“晚籼紫宝”、“香黑米”等常规品种[8-9],也选育出了血黑A、香血粳A、186A等黑米水稻三系不育系[10],实现了杂种优势利用,但关于稻米中总黄酮、花色苷含量的配合力及遗传力的研究较少。伍时照等对2个黑米和2个白米的杂交后代进行研究,结果表明,水稻籽粒种皮的黑(紫)色素沉积是一个由显性基因控制的质量性状,受两对独立遗传的互补基因控制[11]。陈廷文分析了水稻种皮黑色素沉积情况,结果发现,黑色素性状表现受一对显性色素基因控制,存在加性、显性和上位性效应[12]。张名位等采用3个黑米水稻与1个白米水稻配制6个杂交组合,对黑米色素进行遗传分析,结果表明,黑米水稻种皮色素含量的遗传符合加性-显性模型,显性等位基因对提高色素含量起增效作用,隐性等位基因起减效作用,广义遗传力和狭义遗传力均较高[13]。石帮志等对黑米与白米的杂交后代的米色表现进行研究,认为米色性状为数量性状,至少受两对以上显性重叠基因控制,表现有剂量效应[14]。杨海亮采用7×8不完全双列杂交研究了特种红米杂交稻花色苷含量的遗传,结果发现,花色苷含量受亲本一般配合力的影响,受不育系的影响大于恢复系,花色苷含量的广义和狭义遗传率均较高[15]。而王强通过对特种红米杂交稻花色苷含量的遗传分析,结果发现,花色苷的广义遗传力很大,狭义遗传力却很低[16]。王诗文以白米不育系为母本,色稻为父本,采用3×6不完全双列杂交对杂交色稻糙米的花色苷含量进行分析,结果表明,杂交一代花色苷含量偏向于含量低的亲本,母本中一般配合力为正的品种与黑米杂交,其特殊配合力为正值;而母本中一般配合力为负的亲本与黑米杂交,其特殊配合力为负值[17]。孙明茂等报道,水稻花色苷含量是由多基因控制的数量性状,受基因型和环境效应的共同影响[18]。【本研究切入点】本研究所选用的黑米不育系和恢复系,为新选育的黑米稻亲本,株叶形态良好、糙米黝黑、米质优异,是组配黑米两系杂交稻的重要种质资源。目前,尚无针对黑米两系杂交稻亲本的总黄酮、花色苷含量的配合力和遗传力的系统研究,不育系和恢复系在黑米两系杂交稻的总黄酮和花色苷含量性状上所起的作用还有待进一步探明。【拟解决的关键问题】以6个不育系、5个恢复系及其配制的30个杂交组合为材料,通过不完全双列杂交试验,研究黑米两系杂交稻总黄酮、花色苷含量性状的遗传规律,为我国高功能成分黑米两系杂交稻亲本选育、组合测配和评价鉴定等工作提供技术支撑。
1 材料与方法
1.1 试验材料
2016年晚造在福建诏安,以新选育的4个黑米稻两系不育系D18S、D38S、D40S、D43S和2个白米稻两系不育系品272S、品48S为母本,5个黑米两系恢复系R48、R92、R149、R156、R401为父本,按照不完全双列杂交(NCII)设计配制30个黑米两系杂交稻组合。以上材料由福建农林大学农产品品质研究所提供。
1.2 试验方法
2016年11月20日30个黑米杂交组合及其亲本播种于海南三亚,12月20日移栽。试验按随机区组排列,设3个重复,每个小区按8株×8行=64株插秧,株行距20 cm×20 cm,单本插秧,常规管理。每个小区取中间5株测定黑米糙米中总黄酮与花色苷含量,测定方法参考王诗文[17],以小区平均数为计算单位。根据NCII杂交模型进行配合力及遗传力分析,数据分析采用DPS软件分析[19]。
2 结果与分析
2.1 各性状方差和配合力方差分析
供试组合间均方值差异达到极显著水平(表1),说明供试组合间存在遗传差异。进一步进行配合力分析表明,不育系对总黄酮、花色苷含量性状的一般配合力方差均达到极显著水平,表明所配组合的总黄酮和花色苷含量性状中不育系起着重要的作用;恢复系对总黄酮含量性状的一般配合力达到极显著水平,表明在所配组合的总黄酮含量性状中父本起着重要的作用。可见,供试组合的总黄酮、花色含量性状受亲本一般配合力和组合特殊配合力共同决定的,这2个性状的一般配合力和组合特殊配合力的遗传是由加性和非加性基因共同控制的。
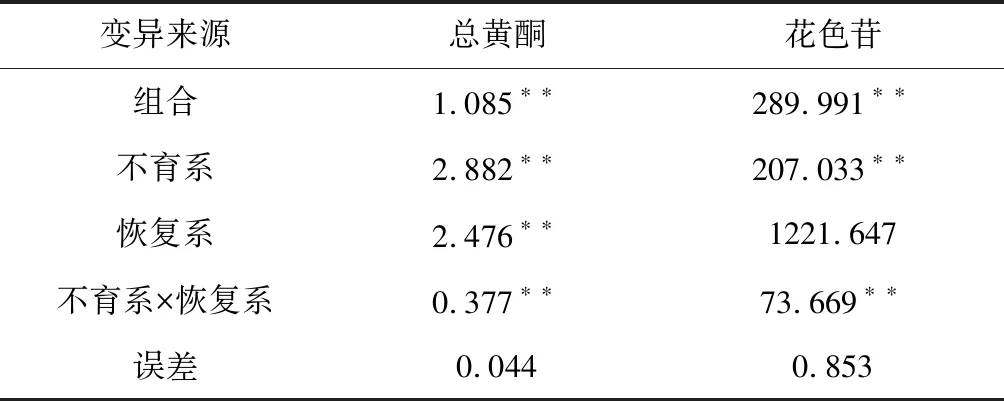
表1 30个组合方差分析及配合力方差分析
注:*、**分别表示P=0.05和P=0.01的显著水平。

表2 亲本各个性状一般配合力效应值
2.2 亲本的一般配合力效应分析
从表2可以看出,供试亲本一般配合力(GCA)效应值在2个性状间存在明显差异,同一性状的各个亲本的GCA也存在明显的差异,表明亲本在不同的性状上的加性效应大小不同,同一性状受到亲本的加性效应也不同。
4个黑米两系不育系D18S、D38S、D40S、D43S在总黄酮和花色苷含量性状上的GCA都有较大的正效应值,D38S的总黄酮和花色苷含量的GCA分别为12.548、51.780,D40S的总黄酮和花色苷含量的GCA分别为9.742、26.595, D38S、D40S在这2个性状上的GCA都有很大的正效应值;黑米恢复系R48的总黄酮和花色苷含量的GCA分别为19.650、17.149,黑米恢复系R92的总黄酮和花色苷含量的GCA分别为2.390、14.225,黑米恢复系R48、R92在这2个性状上GCA都有较大的正效应值。因此,D38S、D40S是总黄酮和花色苷含量性状一般配合力最好的黑米两系不育系,R48、R92是总黄酮和花色苷含量性状一般配合力最好的黑米恢复系。
2.3 各性状特殊配合力效应分析
从表3可以看出,同一性状不同组合间的特殊配合力(SCA)也存在明显差异。30个组合中,总黄酮、花色苷含量性状的SCA变异幅度分别为-16.955~28.741、-39.106~58.746,组合D38S/R92的总黄酮、花色苷含量的SCA效应值分别高达21.675、35.283,组合D40S/R48的总黄酮、花色苷含量的SCA效应值分别高达28.741、58.746,这2个组合是花色苷、总黄酮含量性状SCA效应得到最好的组合。虽然2个白米不育系的一般配合力都很低,但与黑米恢复系杂交,也能组配出特殊配合力较高的杂交组合,组合品48S/R149的特殊配合力比一些2个亲本都是黑米的组合的特殊配合力都要高;2个都是黑米的亲本也可能组配出特殊配合力低的杂交组合,但一般配合力都高的2个黑米亲本所组配的杂交组合D38S/R92、D40S/R48具有更强的杂种优势。因此,在育种中要特别重视一般配合力高的亲本的选择及其选配。

表3 各性状特殊配合力效应值

表4 各性状的基因型方差
注:δ1:母本GCA基因型方差;δ2:父本GCA基因型方差;δ12:父母本互作基因型方差;Vg:GCA基因型方差占总方差比例;Vs:SCA基因型方差占总方差比例;Vg1:母本GCA基因型方差占总方差比例;Vg2:父本GCA基因型方差占总方差比例。

表5 各性状的遗传力
2.4 各性状群体方差和遗传力估计
由表4可以看出,在总黄酮、花色苷含量的遗传中一般配合力所占的比重大,分别为71.53 %、77.57 %(Vg>50 %),表明在这2个性状中,亲本的基因加性效应对杂种一代性状的形成起主导作用。在总黄酮含量中,不育系的一般配合力基因型方差所占的比重为35.85 %,父本为35.67 %,两者差异不大;在花色苷含量中,不育系的一般配合力基因型方差所占的比重为70.72 %比父本6.85 %大,说明在花色苷含量性状上,不育系基因型方差对一般配合力方差贡献更大,在育种过程中要特别注重不育系的选择。
由表5可以看出,这2个性状的狭义遗传力大小顺序为:花色苷>总黄酮。总黄酮含量、花色苷含量的遗传力较高,说明在育种过程中,这2个性状可在早代选择以提高育种效率。
3 讨 论
目前,未见有关稻米中总黄酮含量性状配合力与遗传力的研究,有关花色苷含量性状配合力与遗传力的报道也较少,未见黑米两系杂交稻中关于这两个性状配合力及遗传力的系统研究。
本研究供试组合的总黄酮、花色苷含量性状由亲本一般配合力和组合特殊配合力共同决定的,这两个性状的一般配合力和组合的特殊配合力的遗传由加性和非加性基因共同控制的,这与杨海亮[15]、王强[16]等关于红米中花色苷含量的研究结果相似。黑米花色苷含量性状的遗传中一般配合力所占的比重大,亲本基因的加性效应对这两个性状的形成起主导作用,这与杨海亮研究结果相一致[15],而王强研究认为特殊配合力对红米花色苷影响较大,亲本的非加性效应对红米花色苷含量性状的形成起主导作用[16]。不同人研究结果不一样,可能是由于研究的材料不同造成的。
分析表明,不育系总黄酮含量的一般配合力基因型方差所占的比重与父本相差不大,但不育系花色苷含量的一般配合力基因型方差所占的比重比父本大,不育系基因型方差对花色苷一般配合力方差贡献更大,因此,在育种过程中,要注重不育系对花色苷含量的选择[15]。广义遗传力反映了遗传变异和环境变异的作用,狭义遗传力度量加性遗传效应[20]。总黄酮、花色苷含量的广义遗传力与狭义遗传力都较高,说明这2个性状的加性效应作用突出,受环境影响相对较小,因此,在育种过程中,这2个性状可在早代选择以提高育种效率[21]。
在水稻杂交育种中,性状优良的亲本,所配杂交组合的性状未必优良;性状一般的亲本,所配杂交组合可能有很好的杂种优势,这主要由于亲本间的配合力差异引起的[22]。本研究也存在类似的情况,总黄酮、花色苷含量性状一般配合力高的亲本并不能保证其所配的杂种总黄酮、花色苷含量就高,但一般配合力都高的两个亲本,要获得富含总黄酮、花色苷的杂交组合的概率要更高。
根据GCA和SCA,可以将亲本分成四种类型:第一类,GCA高,SCA大,为最理想的亲本;第二类,GCA高,SCA小,为较好的亲本;第三类,GCA低,SCA大,为有一定的利用价值的亲本;第四类,GCA低,SCA小,为基本无利用价值的亲本[23]。综合黑米不育系和恢复系的配合力表现,可以发现,黑米两系不育系D38S、D40S,黑米恢复系R48、R92在这2个性状的GCA都高,所配组合D38S/R92、D40S/R48的SCA也大,说明D38S、D40S、R48、R92是理想的富集总黄酮和花色苷的黑米两系不育系和恢复系,在生产上有较大的应用前景,通过大量测交组配有望选育出农艺性状、总黄酮和花色苷的特殊配合力好的即高产、高功能成分的强优势组合。好的亲本能够聚集较多的有利基因,两个都是好的亲本进行组配,利用它们加性效应和非加性效应能够配组出优势强的黑米杂交稻新品种,因此,为了提高黑米杂交稻的育种效率,可以在一般配合力高的亲本基础上组配特殊配合力高的黑米杂交稻,同时进行广泛的测交,这样更容易获得强优势的黑米杂交稻组合。
4 结 论
本研究采用6×5不完全双列杂交(NCII)模式对黑米的总黄酮和花色苷含量的性状进行了配合力及遗传力分析。结果表明,总黄酮、花色苷含量性状存在加性效应和非加性效应,亲本基因的加性效应对这2个性状的形成起主导作用,这2个性状遗传力高,可在育种早期世代进行选择以提高育种效率,D38S、D40S、R48、R92是富集总黄酮、花色苷的优异亲本,应用前景较好。
