外源纤维素酶对发酵全混合日粮营养价值、发酵品质和酶活性的动态影响
2019-09-24陈光吉吴佳海尚以顺张蓉何廷章吴彦娄芬熊俊甘小波陈荫琼王普昶韩永芬李世歌舒建虹
陈光吉,吴佳海*,尚以顺,张蓉,何廷章,吴彦,娄芬,熊俊,甘小波,陈荫琼,王普昶,韩永芬,李世歌,舒建虹
(1.贵州省农业科学院草业研究所,贵州 贵阳 550006;2.绥阳县枧坝镇农业服务中心,贵州 绥阳 563307;3.绥阳县草地工作站,贵州 绥阳 563300;4.大方县农业农村局,贵州 大方 551600;5.贵州金农富平生态农牧科技有限公司,贵州 贵阳 550006)
饲草作为反刍动物日粮中最重要的营养物质和能量来源,其干物质中细胞壁比例达40%~65%,在日粮的组成比例达60%~80%,瘤胃微生物可以产生催化水解这些碳水化合物的酶,但其细胞壁中纤维类物质复杂而致密的网状结构限制了反刍动物对它们的消化和利用[1]。研究表明,外源纤维素酶(exogenous fibrolytic enzymes,EFE)添加到日粮中是提高反刍动物纤维利用率的有效途径之一[2-5],这为提高反刍动物纤维利用率,减少“耗粮型”日粮的应用,缓解“人畜争粮”矛盾提供了新的途径[6]。
发酵全混合日粮(fermentation total mixed ration,FTMR)是指根据不同类群或生理阶段反刍动物的营养需要,按设计比例,将青贮、干草等粗饲料切割成一定长度,并和精饲料及各种矿物质、维生素等添加剂进行充分搅拌混合后,装入发酵袋内抽真空或其他方式,创造一个厌氧的发酵环境,经过乳酸发酵的过程,最终调制成的一种营养相对平衡的日粮,其贮存原理与常规青贮饲料类似,即利用乳酸菌发酵后降低饲料pH值,抑制霉菌生长的原理,达到长期保存的目的。与常规的全混合日粮(total mixed ration,TMR)相比,FTMR对反刍动物具有更高的干物质摄入量和瘤胃养分消化率[7]、有效避免长途运输易耗氧腐败[8]和家畜挑食[9]等优势。日本早在20世纪90年代就开发了FTMR技术,而该项技术在我国起步较晚,由于市场需求、配套设备和技术标准等因素,目前仅在北京和上海等大型的奶牛日粮配送中心有所应用,未能得到大范围推广。就日粮配方技术而言,目前的研究主要集中在各地区农副产品[马铃薯(Solanumtuberosum)渣、酱油渣、豆腐渣、果渣等]的发酵品质与有氧稳定性研究和饲草类型替代效应研究[10-11],以及乳酸菌剂[12]、糖蜜、乙酸[13]等添加剂的应用研究,其中大部分添加剂旨在解决前期乳酸菌数量不足导致的养分损失、物料糖分不足导致的发酵失败和有氧条件下的质量稳定性问题。近年来,提高青贮物料自身纤维类物质降解率进而改善发酵品质和营养价值成为青贮添加剂研究的热点之一。这种途径主要围绕木聚糖酶、葡聚糖酶、果胶酶和纤维素酶等非淀粉多糖酶类对青贮物料结构性碳水化合物的降解作用展开,目前主要集中在桑树(Morusalba)青贮[14]、“菌草”青贮[15]和秸秆微贮[16]等单一粗料青贮,且大多表现出对纤维类物质降解的正效应。
然而,针对FTMR的营养成分特性,需要回答关于FTMR中可溶性糖类和缓冲物质含量较高或比例不恰当是否会影响EFE正常酶促反应的问题。为探究这个问题,本试验拟通过研究纤维素酶添加到FTMR中,观察其不同时间点营养价值、发酵品质和酶活性动态变化规律,找出适宜的添加量,为纤维素酶在FTMR中的合理应用提供参考。
1 材料与方法
1.1 试验原料
日粮原料由粗料和精料组成,粗料为第2茬初花期的紫花苜蓿(Medicagosativa)干草和刈割高度为1.8 m的皇草(Pennisetumsinese);精料由玉米(Zeamays)、豆粕、干酒糟及其可溶物(distillers dried grains with solubles, DDGS)、碳酸钙、磷酸氢钙、外源纤维素酶、维生素和微量元素预混料等组成。EFE为纤维素酶和木聚糖酶的复合粉剂,购自湖南尤特尔生化有限公司。试验前,参照Colombatto等[17]的方法测定了外源酶的纤维素酶和木聚糖酶活力:在pH为5.5和39 ℃条件下,纤维素酶和木聚糖酶活力分别为7864.26和3918.47 IU·g-1。
1.2 试验设计
按照《中国肉牛饲养标准》[18]中列出的体重为300 kg,日增重为1 kg的肉牛营养需要的1.1倍营养水平(干物质采食量为7.11 kg·d-1,粗蛋白质摄入量为785 g·d-1)配制基础日粮,按照完全随机试验设计,并根据课题组前期在纤维素酶应用方面的研究基础[19],设置5个纤维素酶添加水平(0、0.4、0.8、1.6、3.2 g·kg-1DM,分别为对照组、EFE+0.4、EFE+0.8、EFE+1.6和EFE+3.2组),分别在发酵7、14、35、49、70 d采集各处理样品进行测定,每个处理的每个时间点分别设置3个重复,日粮配方见表1。
1.3 FTMR制备
2017年8月在贵州省毕节市大方县贵州鸿嘉农牧发展有限公司进行FTMR的制备,实验室分析在贵州省农业科学院草业研究所牧草生物工程实验室进行。预先将EFE添加至精料中按照配方分别单独配制,然后将两种粗料揉切至2 cm左右,按照设计比例(EFE添加量由低到高的顺序进行配制)将精料和粗料投入TMR搅拌机(品牌:银河,型号:TMR5)中均匀混合15 min,每个处理总量为500 kg。搅拌完毕后,运用“四分法”取600 g制作好的TMR料装入真空保鲜袋中(长×宽:30 cm×15 cm),用真空机(品牌:美吉斯,型号:MS1160)抽完袋中空气(真空袋不再向内侧塌陷为止),袋子外贴上标有处理名称等内容的标签,室温贮存待测。
1.4 样品采集和指标测定
贮存后第7、14、35、49、70天开包,按照“四分法”每个处理组采集3份样品(1份400 g,2份100 g):1份400 g样品于65 ℃烘箱中制成风干样,粉碎过40目筛(0.425 mm),用于常规养分和可溶性碳水化合物(water soluble carbohydrate,WSC)含量的测定;2份100 g于-20 ℃保存,待测发酵参数指标和酶活性。
常规养分:参照张丽英[20]的方法测定干物质(dry matter, DM)、粗蛋白质(crud protein, CP)、粗脂肪(ether extract, EE)、粗灰分(crude ash, Ash)、中性洗涤纤维(neutral detergent fiber, NDF)、酸性洗涤纤维(acid detergent fiber, ADF)含量;参考Wang等[21]的方法测定WSC含量;采用PHB-5型便携式pH计(杭州天威工贸有限公司)测定发酵参数中的pH值,采用比色法测定NH3-N含量[22], 二硝基水杨酸(dinitrosalicylic acid,DNS)法测定纤维素酶和木聚糖酶活性(纤维素酶或木聚糖酶活力定义为39 ℃、pH为6.5条件下,在1 min内水解纤维类物质产生相当于1 μg葡萄糖或木糖的酶量,为1个酶活力单位)[23];参考杨智明等[24]测定羧基肽酶和酸性蛋白酶活性[羧基肽酶活力定义为40 ℃、pH为6.5条件下,每kg底物干物质每min释放的游离氨基酸量(μmol),酸性蛋白酶活性定义为在40 ℃、pH 3.0条件下,每kg底物干物质每min水解酪蛋白产生酪氨酸的量(μg)];液相色谱法测定乳酸和挥发性脂肪酸(volatile fatty acid, VFA)含量[25](使用的仪器和色谱条件根据条件摸索结果略作调整),色谱仪器和条件如下:仪器为LC-20A型液相色谱仪[配InertSustainC18(5 μm×4.6 mm×250 nm色谱柱,日本岛津公司],采用外标法进行分析,乙酸、丙酸和丁酸(色谱纯)均购于上海Sigma公司。色谱条件:载气为氦气,流量为2 mL·min-1,分流比为10∶1。采用程序升温,初始温度为120 ℃,以8 ℃·min-1升温至180 ℃,保持5 min。进样口温度为210 ℃,进样量为1 μL。

表1 各处理组日粮配方与营养成分(1)Table 1 Ingredients and nutrient composition of each treatment group(1) (%)
注:(1)为原料组成为新鲜基础,营养成分为干物质基础;(2)为每kg FTMR干物质提供10 mg Cu,50 mg Fe,20 mg Mn,30 mg Zn,0.10 mg Se,0.50 mg I, 0.10 mg Co;(3)为每kg FTMR干物质提供VA 1500 IU, VD 550 IU, VE 10 IU;(4)各处理组日粮营养成分均为实测值。
Note: (1) Ingredient composition with fresh basis and nutrient composition with DM basis; (2) Provided per kg of FTMR: Cu (as copper sulphate)10mg, Fe (as ferrous sulfate) 50 mg, Mn (as manganese sulfate) 20 mg,Zn (as zinc sulfate) 30 mg,Se (as sodium selenite) 0.10 mg, I (as potassium iodide) 0.50 mg,Co (cobalt chloride) 0.10 mg; (3) Provided per kilogram of FTMR : VA 1500 IU, VD 550 IU, VE 10 IU; (4) The nutritional components of each treatment group were measured.
1.5 数据处理
用SPSS 18.0统计软件中的一般线性模型(general linear model, GLM)按照单因子完全随机试验设计进行方差分析,主效应分别为EFE添加水平和发酵时间梯度,用Duncan 法进行多重比较。P<0.05表示差异显著,P<0.10表示有差异显著的趋势。结果用平均值表示,各处理间变异度用平均值的标准误(standard error mistake, SEM)表示。
2 结果与分析
2.1 概略养分和WSC动态变化规律
纵向看,各处理的DM、CP和WSC含量随发酵时间的延长呈显著下降趋势(P<0.05)(表2)。其中,发酵的第14、35和70天的DM含量以对照组最高(P<0.05); 横向看,EFE+0.8和EFE+1.6组的CP含量在发酵的第49天显著高于对照组(P<0.05); 除发酵第0天外,ADF、NDF和WSC含量在其他发酵时间点均以对照组最高(P<0.05),且ADF和NDF含量随酶添加量的提高均存在先下降后上升的趋势,其中EFE+0.8组的ADF含量最低,NDF含量从数值上以EFE+0.4组最低,但与多数EFE添加组差异不显著(P>0.05),各时间点WSC含量以对照组最低。
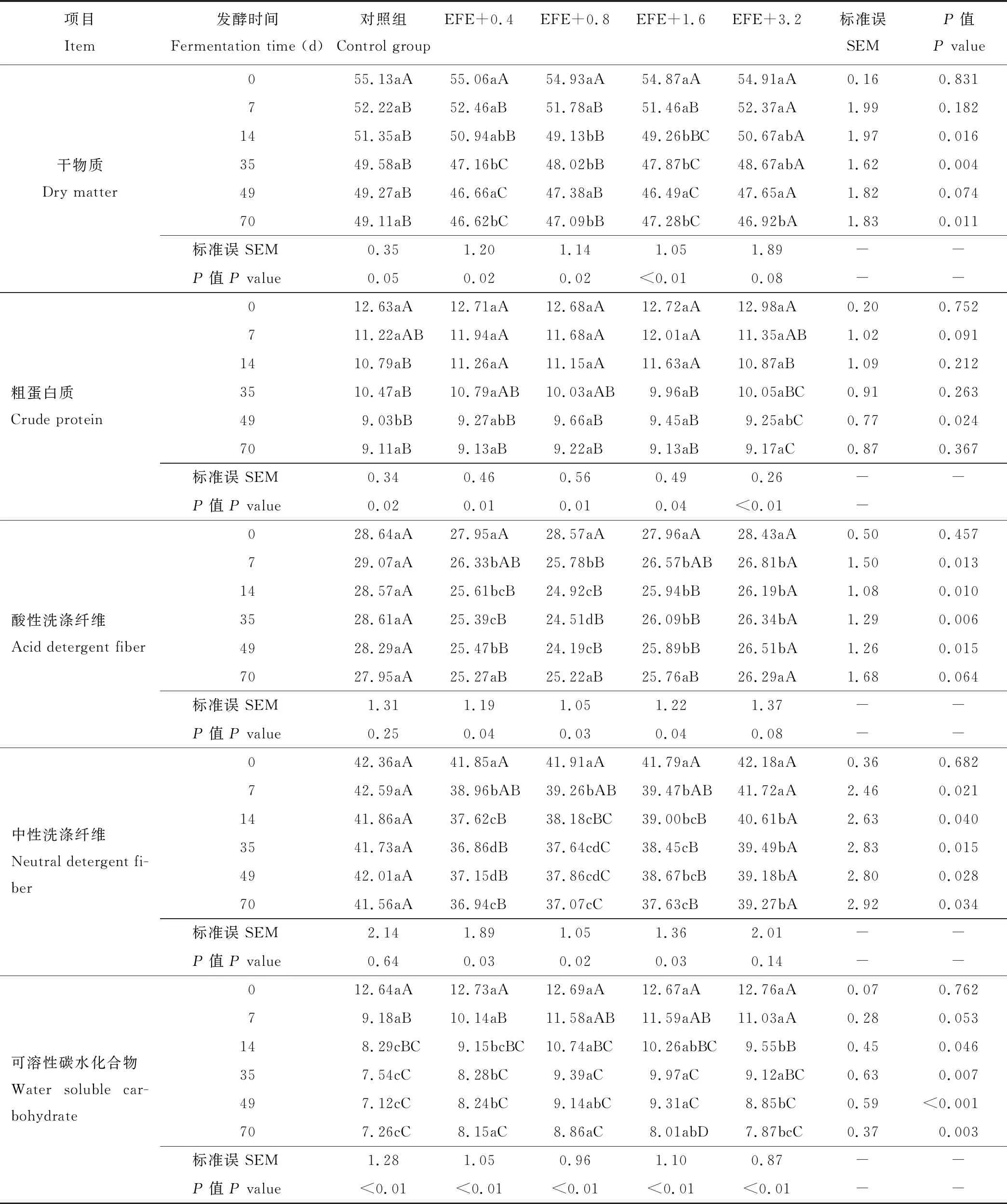
表2 EFE对FTMR 概略养分和WSC含量的动态影响Table 2 Dynamic effects of EFE on FTMR general nutrient and WSC content (%)
注:同行相同小写字母表示组间均值差异不显著(P>0.05),不同小写字母表示差异显著(P<0.05);同列相同大写字母表示组间均值差异不显著(P>0.05),不同大写字母表示差异显著(P<0.05)。下同。
Note: No significant difference with same lowercase letters of the shoulder mark in the mean value between test-groups (P>0.05), and significant difference with different letters (P<0.05); No significant difference with same capital letters of the shoulder mark in the mean value between time-groups (P>0.05), and significant difference with different capital letters (P<0.05). The same below.
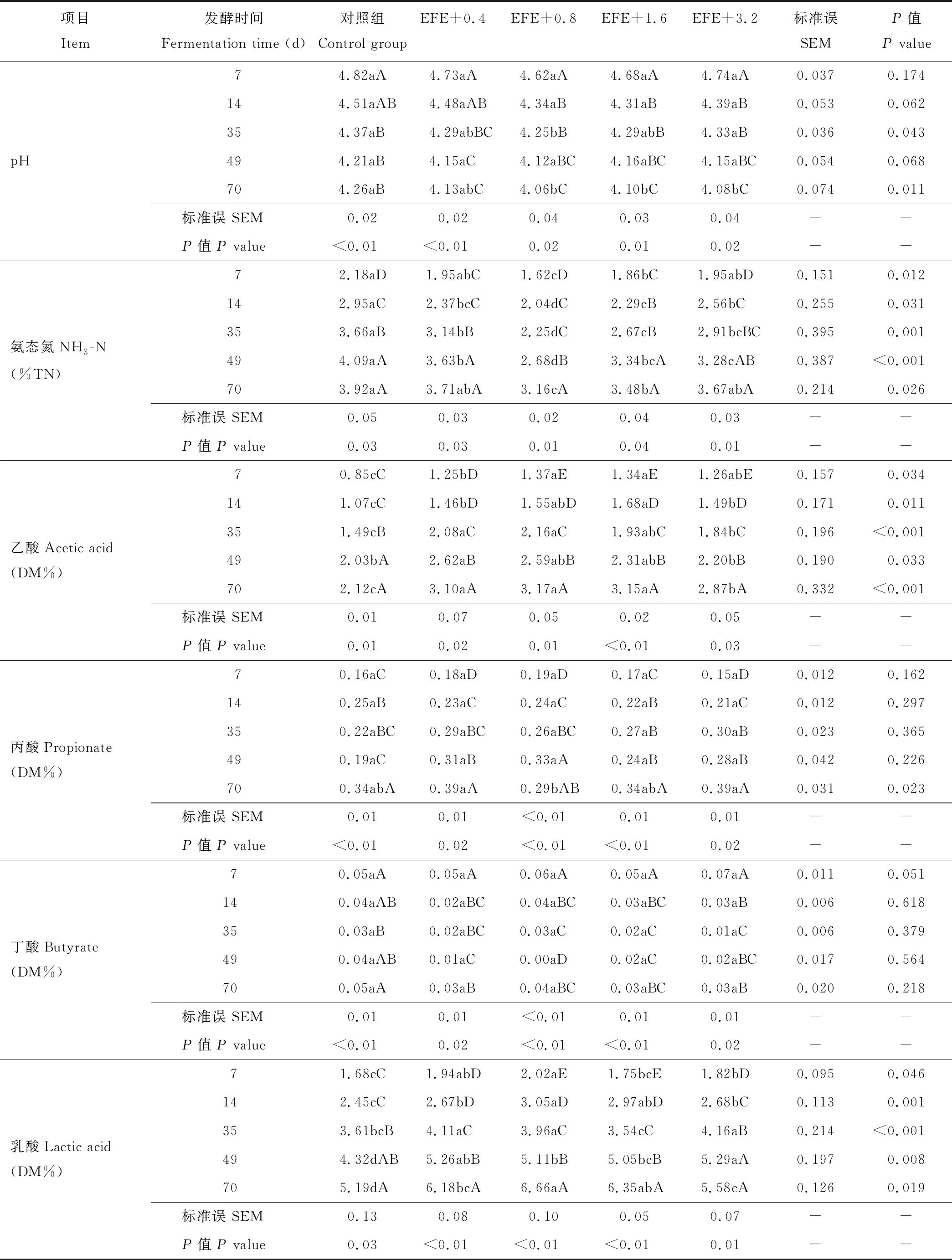
表3 EFE对FTMR发酵参数的动态影响Table 3 Dynamic effects of EFE on FTMR fermentation parameters
2.2 发酵参数动态变化规律
纵向看,随着发酵时间的推移,各处理组pH值均呈现显著下降的趋势(P<0.05);相反,氨态氮、乙酸、丙酸和乳酸含量呈整体上升的趋势(P<0.05),而丁酸含量呈现先下降后上升的趋势(P<0.05)(表3)。横向看,除发酵的第7天,各处理组pH值在其余发酵时间点差异显著(P<0.05)或有差异显著的趋势(P<0.10),且均存在随酶添加量的提高有先下降后上升的趋势,数值上最低点多出现在EFE+0.8组;试验因子对各处理在不同发酵时间点的氨态氮含量有显著影响(P<0.05),且随EFE添加量的提高有先下降后上升的趋势,数值上最低点出现在EFE+0.8组;相反,乙酸和乳酸含量则随酶添加量的提高呈现先上升后下降的趋势,最高点多出现在EFE+0.8组或EFE+1.6组;丙酸含量仅在发酵第70天有显著差异(P<0.05),以EFE+0.8组最低;各处理在各发酵时间点的丁酸含量无显著差异(P>0.10)。
2.3 酶活力动态变化规律
纵向上各处理组纤维素酶、木聚糖酶、羧基肽酶和酸性蛋白酶活力均随发酵时间的推移呈现明显的下降趋势(P<0.05),其中纤维素酶和木聚糖酶活力以对照组下降幅度较大;横向看,试验因子显著影响各处理组的酶活力(P<0.05)(表4);总体规律:随着EFE从0添加量提高到0.8 g·kg-1DM,纤维素酶和木聚糖酶活力多在各发酵时间点呈现上升趋势,但提高到1.6 g·kg-1DM 添加量时则多呈下降趋势,1.6 ~3.2 g·kg-1DM区间时也多呈下降趋势;羧基肽酶和酸性蛋白酶活力在发酵前期(发酵的第7、14、35天)随EFE添加量的提高多呈明显的下降趋势,发酵第35天后呈先下降后上升趋势,且均以对照组的酶活力最大。
3 讨论
3.1 EFE对FTMR概略养分和WSC含量的动态影响
通常,自然条件下青贮饲料中常见的微生物种类包括乳酸菌、酵母菌、好氧细菌、大肠杆菌及其他腐败菌,其中乳酸菌具有嗜酸、糖和盐的特性,其余微生物则主要以糖类和蛋白质为发酵底物,他们是青贮饲料干物质中碳水化合物和蛋白质降解的主要因素,而乳酸菌发酵程度和数量是青贮饲料品质的关键[26]。本试验结果表明,同一处理条件下,随着发酵时间的推移,FTMR的DM、CP含量在发酵的第0~35天的区间呈下降趋势,35 d后DM的损失减少,含量趋于稳定,而全期看CP和WSC含量均呈现发酵时间延长的显著递减效应,这种趋势与张放等[27]和贾婷婷等[28]报道的青贮发酵的养分变化规律一致,表明本试验条件下FTMR在发酵的第35天后微生物菌群活跃度达到高峰期,达到了良好贮存的目的。对于纤维类物质,ADF和NDF含量均随发酵时间的延长呈下降趋势,且以NDF下降幅度较大,主要原因可能是乳酸菌的快速生长导致物料有机酸的迅速积累软化了部分易解离的细胞壁成分,在实际测定时容易被洗涤剂洗脱,但这种解离效应是有限的,NDF组分较多可能是下降幅度较大的主要原因。
研究表明,添加促进发酵型青贮添加剂可通过促进青贮饲料乳酸菌迅速成为优势菌群,减少蛋白质的分解和营养物质的消耗[29-31],外源纤维素酶是其中之一。Nadeau等[32]报道,在苜蓿和鸭茅(Dactylisglomerata)青贮中添加EFE后,其NDF、纤维素和半纤维素降解率分别比对照组提高了33%、37%和27%;此外,牧草品种差异,EFE对NDF的降解程度有所差异,报道指出,黑麦草(Loliumperenne)比对照组提高13%、雀稗草(Paspalumnotatum)提高了21%~33%、小麦(Triticumssp.)秸秆提高了40%~49%、甘蔗(Saccharumofficinarum)渣提高了77%[33-35]。本试验结果显示,EFE添加到FTMR后,同一发酵期内试验组的干物质损失高于对照组,而对照组的氮和WSC含量的损失总体上高于试验组,大部分试验组ADF和NDF降解率高于对照组,且以0.08 g·kg-1DM添加量表现较为好,这与马清河等[36]报道的EFE在青贮饲料中的应用效果基本一致。表明EFE用于高粗料型FTMR中,可降低纤维类物质含量,进而提高能量价值,是配制“节粮型日粮”的可选添加剂之一。
3.2 EFE对FTMR发酵参数的动态影响
pH值是评价青贮饲料发酵品质的最重要的基础指标,底物pH值越低,对有害菌的生长和繁殖的抑制效应越有利,当pH值下降到4.2以下时,能抑制绝大部分有害菌的生长,进而使得青贮饲料很好地保存[37]。氨态氮和有机酸(乙酸、丙酸、丁酸和乳酸等)是微生物为了生长和繁殖的需要,利用底物养分(氮源和碳源)而产生的代谢产物,其中氨态氮和丁酸含量越高表明有害菌发酵程度越深,饲料营养价值降低,乙酸和乳酸含量的迅速提高表征良性发酵,减少营养成分的不良损失,而丙酸与乙酸的相对含量则表示菌群结构的差异导致发酵类型的变化[38]。对于FTMR来说,其发酵原理与青贮饲料基本相同,因此上述评价指标同样适用于FTMR。本试验结果表明,随着发酵时间的推移,各处理组pH值均呈现下降的整体趋势,相反,氨态氮、乙酸、丙酸和乳酸含量呈整体上升的趋势,而丁酸含量呈现先下降后上升的趋势,即发酵品质具有时间效应,这种变化趋势与前人研究结果一致[27,39],数值上看,试验组在发酵第49天的pH值均低于4.20,达到合格青贮饲料的标准,但普遍高于单一原料青贮饲料的pH值[40-41],这可能是由于FTMR本身营养水平较高,特别是含氮量和缓冲能较高,对有机酸有一定的缓冲作用,导致绝对数值高于单一青贮饲料,因此FTMR中的可溶性碳水化合物含量与含氮量或缓冲能之间的互作效应可能是未来需要研究和回答的问题,这为兼顾FTMR配方营养水平合理性和发酵品质的优良性提供理论依据。
本试验中,氨态氮与总氮的比值随EFE添加量的提高有先上升后下降的趋势,且数值均小于10%,达到MacDonald[42]报道的保存良好的青贮饲料的标准,发酵正常。同时,表明EFE在抑制FTMR蛋白质的降解具有剂量效应,这种效应在乙酸和乳酸含量的变化规律中得到对应体现,这结果与pH值结果相互印证,且以0.8和1.6 g·kg-1DM的添加效应较好。剂量效应的原因可能是由于过高剂量的EFE吸附和占据了有限的植物细胞壁中纤维素晶体的大部分结合域,竞争性地抑制了FTMR发酵过程中微生物自身产生的纤维类物质降解酶的作用效应[19]。这种效应与Nsereko等[43]报道的关于纤维素酶与反刍动物瘤胃纤维降解类微生物的关系的研究结果类似[44]。
从作用途径和机理上看,任海伟等[45]利用高通量测序技术研究了EFE对青贮饲料细菌多样性的影响,结果表明,EFE通过对底物发酵前期部分结构性碳水化合物的降解,补充了乳酸菌可利用的可溶性糖类,一方面让乳酸菌迅速成为优势菌群,另一方面乳酸的迅速积累又加速抑制了芽孢杆菌、耶尔森氏菌、沙门氏菌、弯曲杆菌和肠杆菌等致病腐败菌,本试验中各处理的乳酸含量变化趋势也印证了这个结果,同时也解释了EFE减少了底物氮的损失规律。
3.3 EFE对FTMR酶活力的动态影响
纤维素酶是由内切葡聚糖酶(endo-1,4-β-D-glucanase)、外切葡聚糖酶(exo-1,4-β-D-glucanase)和β-葡萄糖苷酶(β-1,4-glucosidase)组成的酶类总称,其作用与由木聚糖组成的半纤维素酶具有协同效应,它们是自然界水解纤维类物质的主要形式[46]。关于青贮饲料中应用纤维素酶复合物的研究,前人已有报道[47-48],均表现出对青贮饲料发酵品质和营养价值方面的正效应,但还未见从发酵后酶活力的变化规律探究其作用机理的报道。本研究结果表明,各处理组纤维素酶和木聚糖酶活力随发酵时间的延长呈下降趋势,表明随着发酵环境有机酸的积累,pH下降对酶活力产生了抑制作用,这与酶促反应条件的规律和本试验中NDF和ADF的降解率结果相吻合。研究表明,纤维素酶的来源决定了其酶促反应的作用条件,其中霉菌产生酸性和中性纤维素酶,细菌则产生碱性或酸性纤维素酶;酸性纤维素酶在酸性条件下较稳定,其反应最佳的pH值范围为 3~6;中性纤维素酶在中性条件下较稳定,其反应最佳期的pH值为6~7;碱性纤维素酶的最佳反应pH值为9~10[49]。本试验结果显示,同一发酵时间点随EFE添加量提高纤维素酶活力呈明显的上升趋势,并未随pH值降低而显著下降,这可能与本试验所用的EFE属酸性纤维素酶类型有关,但具体原因还需要试验研究进一步证明。
研究证实,羧基肽酶和酸性蛋白酶是牧草中存在的两种主要的植物蛋白水解酶,植物脱离土壤或其他生存条件后,自身代谢加快后会激活更多的蛋白酶将植物蛋白质水解为小肽、氨基酸和氨态氮[50],因此这两种酶的活力可以代表青贮饲料中蛋白降解的能力,以及潜在的有氧稳定性。本试验结果表明,羧基肽酶和酸性蛋白酶活力随发酵时间延长而逐渐降低,且以羧基肽酶降幅较大,与杨智明等[24]报道的苜蓿青贮中酶活力变化规律一致,且pH值的变化规律基本吻合。此外,本试验结果还可看出,EFE有加速两种蛋白酶活力降低的作用,但其机理是通过降低有机酸积累,间接抑制其酶活力还是通过直接的途径需要进一步研究证实。
4 结论
本试验条件下,FTMR的营养价值和发酵品质随发酵时间呈动态变化;添加EFE可促进FTMR中纤维类物质的降解、有机酸的积累和自身纤维类物质的酶活力,抑制羧基肽酶活力和酸性蛋白酶活力,减少蛋白质的损失,且存在添加剂量效应,适宜添加量为0.8~1.6 g·kg-1DM。
