一个分离自贵州省茶叶上的拟盘多毛孢属新种
2019-09-13陈立杰陈通政
陈立杰, 宋 玉, 陈通政, 刘 颖, 王 勇
(贵州大学, 贵阳 550025)
拟盘多毛孢属(Pestalotiopsis)按照最新的分类系统属于子囊菌门,盘菌亚门,粪壳菌纲,炭角菌亚纲,黑盘孢目,被毛孢科。该真菌类群地球上普遍存在,尤其在热带和亚热带生物多样性特别丰富[1],此外,该真菌类群不具有很高的专一性宿主,因此往往能引起许多植物的不同病害症状,造成不同程度的减产[2-10]。根据前人研究[4],Pestalotiopsis分类的主要形态学指标涵盖:第一,分生孢子中间有色孢的颜色特点(包括同色,异色,橄榄绿色及褐色);第二,孢子大小范围,形状特点及分隔数目;第三,孢子附属丝(顶端)的特点、基部附属丝的有无及数目;第四,产孢器(分生孢子盘)、分生孢子梗及产孢细胞结构的特点等[4]。近年来,随着分子生物学技术的发展,在真菌类群的分类学研究中,分子系统学技术(基于DNA序列的碱基差异的分析)发挥着越来越重要的作用且优势明显。本研究结合形态学和分子系统学的方法对分离自贵州茶叶上的拟盘多毛孢真菌进行分类学评价。
1 材料与方法
1.1 采集标本
采集地:花溪区黔陶乡赵司村茶场;品种:福鼎大白茶;采集时间:2011年11月28日;标本及所获菌株均保存于贵州大学农学院植物病理教研室标本室。
1.2 分离纯化及保存菌种
菌种的分离纯化及保存方法参照宋玉[11]的方法。
1.3 形态学描述
所分离获得的菌株在25 ℃条件下培养7~10 d,等到肉眼观察有大量油滴状黑色或深褐色颗粒(分生孢子盘及分生孢子)出现后在体视镜下观察菌落形态并挑取分生孢子及分生孢子梗等制作玻片,显微镜(奥林帕斯BX 53)在不同放大倍数下(20×,40×,100×)观察真菌的典型结构特征并拍摄分生孢子、产孢细胞和分生孢子盘的高清图片,采用PhotoShop软件选择典型的照片组成合格的图版,Image Frame Work软件测量拟盘多毛孢菌不同繁殖结构的形态学指标并完成相应的形态学描述。
1.4 DNA的提取、PCR扩增及测序
1.4.1基因组DNA的提取
采用DNA快速提取试剂盒提取拟盘多毛孢菌的基因组DNA,具体步骤参考宋玉[11]的方法。
1.4.2PCR反应体系,引物及扩增程序
1) 反应体系。
PCR总反应体系为25μL:不同基因正反向引物分别1μL,Template DNA 1μL,双蒸水 9.5μL,2倍的PCR Master Mix(2×Taq DNA聚合酶,2×PCR 缓冲液以及2×dNTP)共计12.5μL[11]。
2) 不同基因引物序列及PCR程序。

注:模式标本为HGUP 4086,其中A~H为分生孢子;I~J为分生孢子梗和产孢细胞。标尺:A~J=20 μm。图1 中国茶拟盘多毛孢
ITS 基因DNA片段:
通用引物(ITS 4和ITS 5),引物序列见参考文献[12]。
反应程序:95 ℃下总预变性0.5 min,然后95 ℃变性反应0.5 min,52 ℃退火反应45 s,72 ℃延伸1.5 min,反应进行35个循环后72 ℃总延伸10 min[11],反应完成后4 ℃下保存或及时取出。
β-tubulin(β-微管蛋白)基因DNA片段:
特异引物Bt2a和Bt2B,引物序列见参考文献[13-14]。
反应程序:93 ℃总预变性3 min,然后95 ℃变性1 min,55 ℃退火50 s,72 ℃延伸1 min,反应进行35个循环后72 ℃总延伸10 min[14-15],反应完成后4 ℃下保存或及时取出。
tef1(延伸因子)基因DNA片段:
特异引物EF 1-526F和EF 1-1567 R,引物序列见参考文献[16]。
反应程序:94 ℃总预变性5 min,然后94 ℃变性0.5 min,63到53 ℃或66到56 ℃(每循环降低1 ℃)退火50 s,共10个循环;94 ℃变性0.5 min,63或66 ℃退火55 s, 72 ℃延伸1.5 min,反应进行36个循环后72 ℃总延伸7 min[16],反应完成后4 ℃下保存或及时取出。
1.5 不同基因的DNA序列分析
将编辑好的三个基因(ITS+β-tubulin+tef1)序列加合文件在PAUP* 4.0 b 10[17]中运行,算法设定为最大简约法(Maximum Parsimony,MP)。采用启发式搜索(heuristic search)算法并应用树二等分再连接选项(Tree Bisection-reconnection,TBR)来进行分枝交换,自举法(Bootstrap)检测设置为1000次。程序运行完成后所生成的系统发育树的拓扑结构在Treeview软件中显示,选择最优树的拓扑结构在Figtree软件中进行编辑。
2 结果与分析
2.1 形态学特征描述
中国茶拟盘多毛孢(Pestalotiopsiscamellia-sinensisYong Wang bis, Y. Song, & K.D. Hyde, sp. nov.)拟盘多毛孢属真菌子实体球形,黑色,未成熟前往往寄生于植物组织表皮下方,直到成熟后方外露,呈现凸起(黑色的小颗粒)。分生孢子梗很短至不明显,产孢细胞呈现安培瓶状。分生孢子梭形或近梭形,偶有弯曲,具4个横隔膜(5个细胞),大小范围是27~35 × 7~10μm (平均值为31.5 × 8.5μm); 其中中间的3个深色胞均为浅褐色,长度为17~23μm (平均值为20μm),分隔处稍缢缩,横膈膜颜色有加深,分生孢子的壁不光滑具点突;中间深色3细胞自下而上第一个其长度是5~8μm (平均值为6.5μm),第二个细胞的长度为6~8μm (平均值为7.4μm),第三个细胞的长度为5~8μm (平均值为7μm);中间3细胞两端的细胞近透明,亚圆锥形,长度为5~8μm (平均值为6.5μm);其中顶端细胞具4~6根管状的附属丝,附属丝往往具有分枝,长度为20~40μm (平均值为30μm);分生孢子的基细胞略钝圆,附属丝往往缺失。
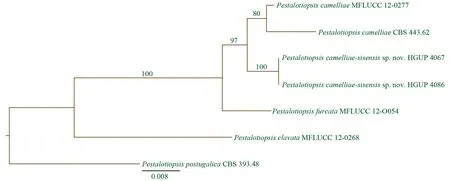
图2 中国茶拟盘多毛孢的DNA序列分析
模式标本:贵州省贵阳市花溪区黔陶乡赵司村,茶树(福鼎大白茶)叶片,模式标本编号HGUP 4086,采集人:宋玉,采集时间:2011-11-28。
其他标本:贵州省贵阳市花溪区黔陶乡赵司村,茶树(福鼎大白茶)叶片,标本编号HGUP 4076,采集人:宋玉,采集时间:2011-11-28。
2.2 不同基因的DNA序列分析
通过ITS,β-微管蛋白以及延伸因子DNA序列构建的系统进化树,我们发现中国茶拟盘多毛孢与山茶拟盘多毛孢(PestalotiopsiscamelliaeY.M. Zhang, Maharachch. & K.D. Hyde)和顶生分枝拟盘多毛孢(PestalotiopsisfurcataMaharachch. & K.D. Hyde)保持了比较近的遗传进化关系,同时又保持了相对独立的分类地位(形成3个进化分枝),分别获得100%及97%的自举支持率。就形态学而言,三者也存在较明显的差别。中国茶拟盘多毛孢的分生孢子大小比顶生分枝拟盘多毛孢的小,但比山茶拟盘多毛孢的大。此外,中国茶拟盘多毛孢顶端附属丝的数目也与其它2个种的也不相同,并且顶端附属丝普遍会分枝,但近似种没有表现出这些形态特征特征[6,17]。因此,结合以上DNA序列分析和形态学特征的研究,我们将中国茶拟盘多毛孢确定为拟盘多毛孢属的一个新种。
3 讨 论
中国茶拟盘多毛孢,山茶拟盘多毛孢和顶生分枝拟盘多毛孢这三个种均分离自茶树上,其中山茶拟盘多毛孢分离自云南,顶生分枝拟盘多毛孢分离自泰国,而本研究中的新种来自贵州省[6,17]。此外,山茶拟盘多毛孢和顶生分枝拟盘多毛孢均可以引起茶树的灰斑病,而我们这个种还没有引起病害的直接证据。尽管形态学和分子系统学的结论已经证明我们所发现的新种与山茶拟盘多毛孢和顶生分枝拟盘多毛孢不同,然而我们不能掉以轻心,贵州省作为茶叶之乡且种植面积全国第一[18],应该时刻关注病害的发生动态,因此我们在后续研究中会探究引起病害的可能性及危害性。
