重组自交系稻米品质性状的相关性分析及QTL定位
2019-09-10秦钢夏秀忠粟学俊陈彩虹梁曼玲阎勇
秦钢 夏秀忠 粟学俊 陈彩虹 梁曼玲 阎勇

摘要:【目的】分析重組自交系稻米品质性状的相关性,并对其进行数量性状座位(QTL)定位,为水稻品质性状遗传改良提供理论依据。【方法】以珍汕97B、柳沙油占202及二者杂交构建的174个重组自交系群体为材料,对12个稻米品质性状进行相关性分析和QTL定位。【结果】3个稻米碾磨品质性状(出糙率、精米率和整精米率)间均呈极显著正相关(P<0.01,下同)。6个稻米外观品质性状中,除粒长与垩白率、垩白度和透明度间无显著相关性(P>0.05)外,其他性状间均呈极显著正或负相关。3个稻米蒸煮食味品质性状中,胶稠度与直链淀粉含量和碱消值呈极显著负相关。碾磨品质性状、外观品质性状和蒸煮食味品质性状间也存在不同程度相关性,如垩白粒率和垩白度与除粒长外的其他9个性状呈显著(P<0.05,下同)或极显著相关等。在第5和第6染色体上共检测出出糙率、精米率、粒宽、长宽比、垩白率、垩白度、直链淀粉含量、胶稠度和碱消值9个性状的11个QTL,其中9个QTL的表型变异贡献率大于10%,为主效QTL,且显著或极显著相关性状的QTL定位区域存在趋同现象。【结论】重组自交系群体12个稻米品质性状存在互作效应,与QTL定位结果相互印证。鉴定所得主效QTL可用于水稻稻米品质相关性状的遗传改良。
关键词: 水稻;重组自交系;品质性状;相关性分析;QTL定位
中图分类号: S511.035.3 文献标志码: A 文章编号:2095-1191(2019)04-0703-07
Abstract:【Objective】In the present study,correlation analysis of rice recombinant inbred line was studied, and quantitative trait loci mapping was carried out to provide reference for rice quality traits genetic improvement. 【Method】 One hundred and seventy-four recombinant inbred line(RIL) populations derived from the cross between Zhenshan 97B and Liushayouzhan 202 were used to conduct correlation analysis and QTL mapping of rice quality traits. 【Result】The results showed that three traits of milling, namely percentage of brown rice,milled rice rate and head rice rate had extremely significant correlation between each other(P<0.01, the same below). Among the six appearance traits, except that grain length had no significant correlation with chalky grain rate, chalkiness and transparency(P>0.05), the other traits had extremely positive correlation or extremely negative correlation between one and another. In the three cooking and eating quality traits, gel consistency had extremely negative correlation with amylase content and gelatinization temperature. Milling quality traits, appearance quality traits, cooking and eating quality traits also had correlation at various degrees. For example, chalky grain rate and chalkiness was significantly(P<0.05, the same below) or extremely correlated with all the traits except grain length. For the nine traits(percentage of brown rice,milled rice rate,grain width,ratio of length to width,chalky grain rate,chalkiness,amylase content,gel consistency and gelatinization temperature),eleven QTLs were identified locating on chromosome 5 and chromosome 6. There were nine QTLs whose phenotypic variation contribution rates were over 10%, they were major effects QTLs. The QTLs location area of the traits which had significant or extremely significant correlation between each other were apt to be located in the same region. 【Conclusion】There are interaction effects among the 12 grain quality-related traits tested on recombinant inbred line populations. Such effects can be testified in the further QTL mapping detecting. Major QTLs detected in the present study can be used in genetic improvement of rice quality-related traits.
Key words: rice; recombinant inbred line; quality-related traits; correlation analysis; QTL mapping
0 引言
【研究意义】水稻是我国最重要的粮食作物,受我国人口压力和耕地面积制约,高产一直是我国水稻育种专家追求的首要目标(蔡薇等,2017)。随着生活水平的不断提高,人们对稻米品质提出更高的要求,优质逐渐成为与高产并重的育种目标。稻米品质主要涉及碾磨加工、外观、蒸煮食味和营养等品质(黄发松等,1998),其中,碾磨加工品质性状主要包括出糙率、精米率和整精米率;外观品质性状主要包括米粒长、米粒宽、长宽比、垩白率、垩白度和透明度;蒸煮食味品质性状主要包括直链淀粉含量、胶稠度和碱消值;营养品质性状主要包括稻米中的蛋白质含量和氨基酸组成(张昌泉等,2016)。因此,鉴定发掘稻米品质性状的数量性状座位(QTL)并了解各性状间的互作效应对科学制定有效的品质改良育种方案具有重要意义。【前人研究进展】目前,大量关于稻米出糙率、精米率和整精米率等碾磨加工品质性状的QTL已完成分子标记定位。梅捍卫等(2002)利用Lemont和特青杂交所得重组自交系检测出1个精米率和4个整精米率的主效QTL。梅德勇等(2012)利用特青和IRBB杂交所得重组自交系检测出8个糙米率、2个精米率和2个整精米率的主效QTL。刘家富等(2007)利用特青和云南元江野生稻构建一个渗入系,并检测出3个糙米率和1个整精米率的QTL。穆平等(2007)利用IRAT109和越富的加倍单倍体群体检测出1个糙米率、1个精米率和3个整精米率的主效QTL。胡霞等(2011)利用测258和IR75860的导入系检测出4个糙米率、5个精米率和6个整精米率的QTL。关于外观品质性状,GS3、GW7、GW2、GW5和GS5等粒长或粒宽的基因已被克隆,其中GW7/GL7基因编码TONNEAU1募集基序蛋白,控制粒长(Wang et al.,2015);GW2基因编码E3泛素连接酶活性环蛋白,控制粒宽(Song et al.,2007);GW5基因编码富含精氨酸蛋白,控制粒宽(Weng et al.,2008;方先文等,2017);GS5基因编码氨酸羧肽酶,也控制粒宽(Li et al.,2011)。与控制粒长或粒宽基因克隆相比,对垩白相关性状的研究进展不明显,尚停留在QTLs分子定位水平。如Zhou等(2009)利用培矮64S和9311构建的染色体片段置换系对垩白相关性状QTL进行精细定位,结果发现,垩白粒率QTL qPGWC-7定位在44 kb的区段内;Guo等(2011)利用Asominori和IR24构建的染色体片段代换系对垩白相关性状QTL进行精细定位,结果发现,垩白粒率QTL qPGWC-8定位在140 kb的区段内。关于蒸煮食味品质性状,其被大量基因所调控,且分子机理也得到较深入的研究,如直链淀粉含量由Wx基因调控(Wang et al.,1995),碱消值的主效基因是alk(高振宇等,2003),胶稠度受Wx基因和其他微效基因调控(高维维等,2017)。较多学者对蒸煮食味品质性状进行相关性分析和QTL定位分析,结果发现大量相关调控基因或主效QTL,且各性状间存在互作效应(邵高能等,2009;晁园等,2012;梅德勇等,2012;彭强等,2016)。邵高能等(2009)利用重组自交系检测出直链淀粉含量、胶稠度和碱消值的主效QTL各一个,分别为qAC6、qGC6和qASV6,其中qASV6位于第6染色体短臂Wx基因区域。晁园等(2012)利用重组自交系检测到5个糊化温度QTL、3个直链淀粉含量QTL,其中直链淀粉含量主效QTL qAC6位于Wx基因处,而糊化温度主效QTL qASV6-4和qASV6-5也位于第6染色体的alk基因处。【本研究切入点】至今,鲜见同时对稻米碾磨加工、外观和蒸煮食味等相关品质性状进行相关性分析和QTL定位的研究报道。【拟解决的关键问题】以珍汕97B、柳沙油占202及二者杂交构建的重组自交系为材料,对其12个稻米品质性状进行相关性分析及QTL定位,综合研究品质性状间的互作效应,以期从分子水平揭示各性状形成的遗传机制,为水稻的品质改良提供理论依据。
1 材料与方法
1. 1 试验材料
供试水稻材料为柳沙油占202、珍汕97B及二者杂交构建的174个重组自交系(F6代),由广西农业科学院水稻研究所提供。Ex Taq DNA聚合酶、DL2000 DNA Marker购自宝生物工程(大连)有限公司。主要仪器设备:PCR仪(ABIVeriti FASTPCR仪)。
1. 2 田间试验和性状考查
田间试验于2016年早造在广西农业科学院试验田进行,174个重组自交系及其亲本柳沙油占202和珍汕97B按照2重复完全随机试验方案种植。同一重复中每个株系种1小区,每小区3行,每行15株,正常田間管理。水稻成熟期,每个小区随机选取20株混合收种,室内自然通风晾干后,在常温下储藏3个月,按照中华人民共和国农业部标准米质测定方法,考察以下性状:出糙率、精米率、整精米率、粒长、粒宽、长宽比、垩白率、垩白度、透明度、直链淀粉含量、胶稠度和碱消值。各品质性状取重复的平均值作为分析数据进行QTL定位及相关性分析。
1. 3 遗传图构建和QTL定位
分蘖盛期取新鲜叶片迅速进行液氮固定,-80 ℃保存备用,采用CTAB法抽提其DNA。具体抽提步骤、SSR扩增反应体系、扩增程序及电泳参数设置参照张月雄等(2018)的方法。从496个SSR标记中筛选获得56个在亲本间具有较好多态性的构图标记,用JoinMap 4.0构建遗传连锁图。该遗传图谱覆盖水稻基因组664.9 cM,标记间的平均遗传距离为11.8 cM。采用MAPQTL 6.0的区间作图(Internal Mapping)方法进行品质性状的QTL分析,扫描步长设定为默认值(1.0 cM),LOD值设定为3.0,计算出的LOD值大于3.0将被认为存在假定的QTL,同时计算各QTL对各性状的表型变异贡献率和加性效应。
1. 4 统计分析
利用Minitab进行正态性分布检测和相关性分析等数理统计。
2 结果与分析
2. 1 亲本及重组自交系群体的稻米品质性状表现
由表1可知,柳沙油占202的碾磨加工品质、外观品质和蒸煮食味品质均高于珍汕97B;重组自交系群体中,除粒宽、长宽比和透明度外的其他外观品质性状均存在超亲分离现象,碾磨加工品质和蒸煮食味品质性状也均出现双超亲分离现象。重组自交系群体的出糙率、粒长、粒宽和长宽比符合正态分布。可见,大部分稻米品质性状出现双超亲分离,符合数量性状遗传分布特征。
2. 2 重组自交系群体稻米品质性状的相关性分析结果
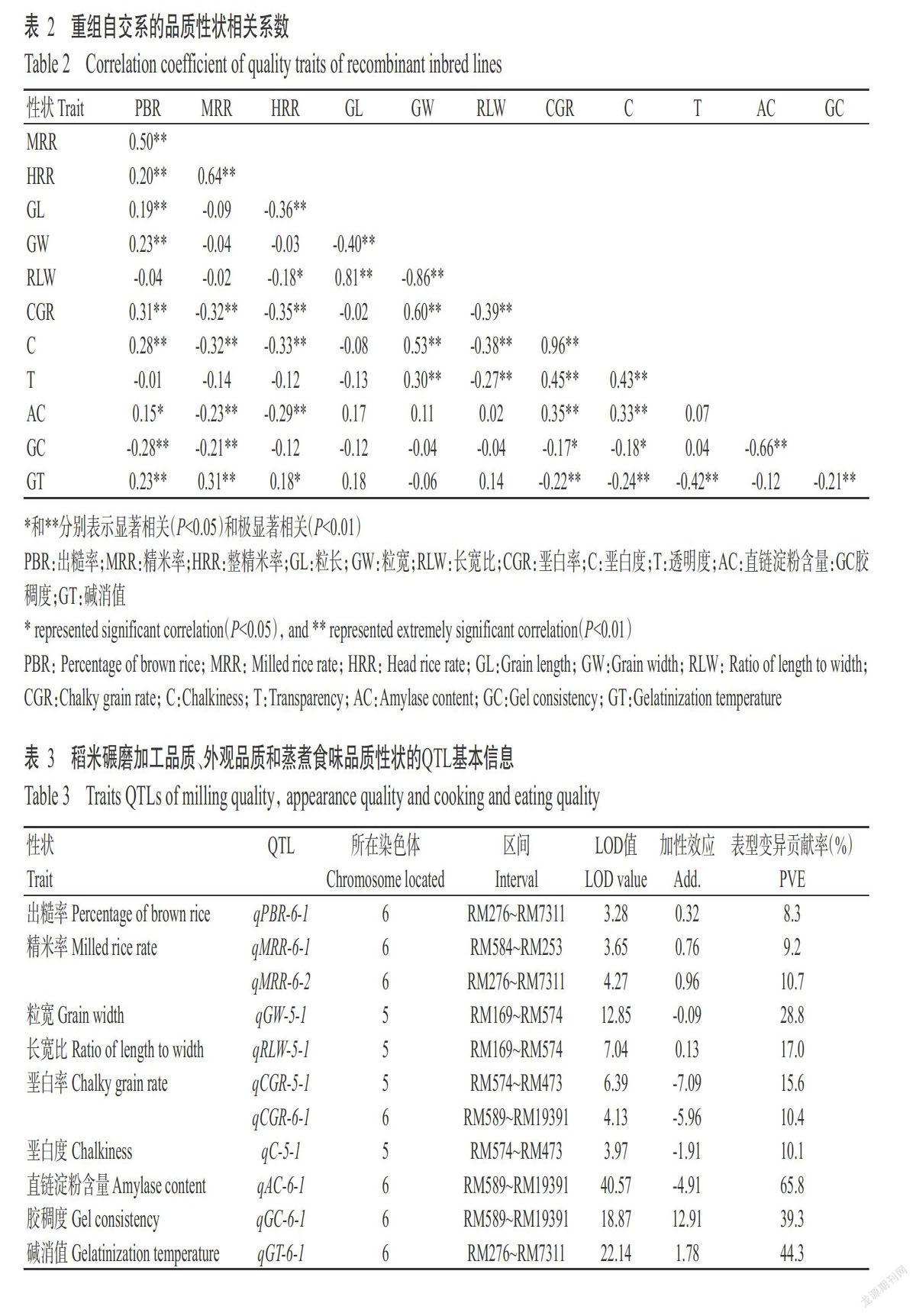
重组自交系群体的碾磨加工品质性状、外观品质性状和蒸煮食味品质性状间的相关性分析结果如表2所示。3个碾磨品质性状间均呈极显著正相关(P<0.01,下同),且与外观品质性状和蒸煮食味品质存在不同程度相关性,其中,出糙率与粒长、粒宽、垩白率、垩白度和碱消值呈极显著正相关,与直链淀粉含量呈显著正相关(P<0.05,下同),与胶稠度呈极显著负相关;精米率与垩白率、垩白度、直链淀粉含量和胶稠度呈极显著负相关,但与碱消值呈极显著正相关;整精米率与粒长、垩白率、垩白度和直链淀粉含量呈极显著负相关,与长宽比呈显著负相关,但与碱消值呈显著正相关。
外观品质性状中,除粒长与垩白率、垩白度和透明度间无显著相关性外,其他性状间均呈极显著正或负相关,其中粒长与粒宽呈极显著负相关,与长宽比呈极显著正相关;粒宽与长宽比呈极显著负相关,但与垩白率、垩白度和透明度呈极显著正相关;长宽比与垩白率、垩白度和透明度呈极显著负相关;垩白率、垩白度和透明度3个性状间呈极显著正相关。垩白粒率和垩白度均与除粒长外的其他9个稻米品质性状呈显著或极显著相关。
蒸煮食味品质性状中,胶稠度与直链淀粉含量和碱消值呈极显著负相关。蒸煮食味品质性状与外观品质性状存在不同程度相关性,其中,直链淀粉含量与垩白率和垩白度呈极显著正相关,胶稠度与垩白率和垩白度呈显著负相关,碱消值与垩白率、垩白度和透明度呈极显著负相关。
2. 3 碾磨加工品质性状的QTL定位结果
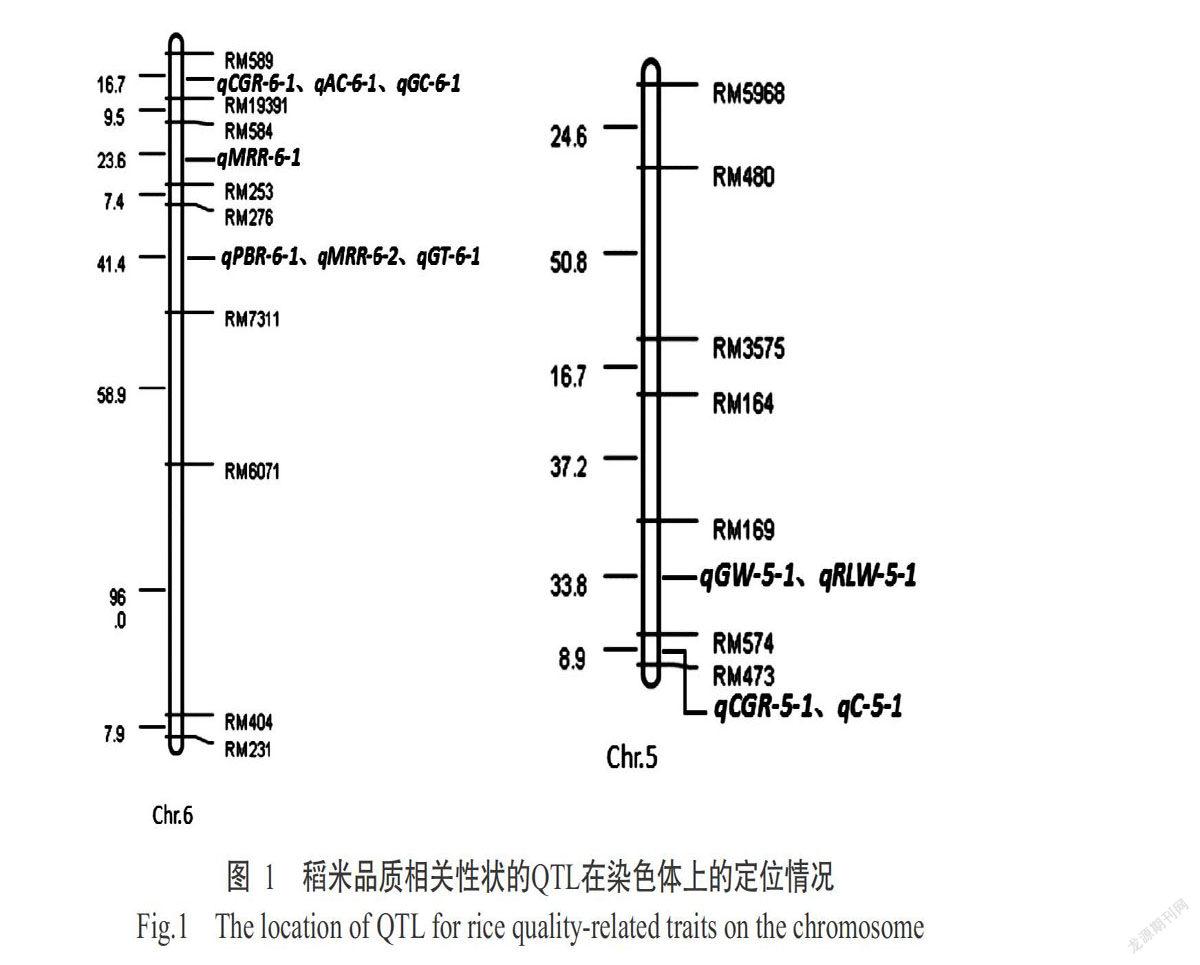
如表3和图1所示,在第6染色体共检测出3个碾磨加工品质性状的QTL,包括1个出糙率QTL(qPBR-6-1)和2个精米率QTL(qMRR-6-1和qMRR-6-2),其中,qPBR-6-1位于RM276和RM7311标记之间,加性效应为0.32,表型变异贡献率为8.3%;qMRR-6-1位于RM584和RM253之间,加性效应为0.76,表型变异贡献率为9.2%;qMRR-6-2位于RM276和RM7311之间,加性效应为0.96,表型变异贡献率为10.7%,为主效QTL。可见,出糙率QTL qPBR-6-1和精米率QTL qMRR-6-1共同位于RM276和RM7311标记之间,且上述相关性分析结果(表2)显示,出糙率与精米率呈极显著正相关,表明这两个性状存在共同的内在分子遗传机制,以支撑表型的高度相关性。
2. 4 外观品质性状QTL定位结果
由表3和图1可知,在第5和第6染色体共检测出5个外观品质性状的QTL,包括1个粒宽QTL(qGW-5-1)、1个长宽比QTL(qRLW-5-1)、2个垩白率QTL(qCGR-5-1和qCGR-6-1)和1个垩白度QTL(qC-5-1),其中,qGW-5-1位于第5染色体RM169和RM574标记之间,加性效应为-0.09,表型变异贡献率为28.8%,为主效QTL;qRLW-5-1也位于第5染色体RM169和RM574标记之间,加性效应为0.13,表型变异贡献率为17.0%,为主效QTL;qCGR-5-1位于第5染色体RM574和RM473标记之间,加性效应为-7.09,表型变异贡献率为15.6%,为主效QTL;qCGR-6-1位于第6染色体RM589和RM19391标记之间,加性效应为-5.96,表型变异贡献率为10.4%,为主效QTL;qC-5-1位于第5染色体RM574和RM473标记之间,加性效应为-1.91,表型变异贡献率为10.1%,为主效QTL。综上所述,粒宽、垩白率和垩白度QTL的加性效应均为负数,说明增效等位基因来自珍汕97B;粒宽QTL qGW-5-1和长宽比QTL qRLW-5-1均位于第5染色体RM169和RM574之间,垩白率QTL qCGR-5-1和垩白度QTL qC-5-1均位于第5染色体RM574和RM473标记之间,且上述相关性分析结果(表2)显示,粒宽与长宽比、垩白率和垩白度均呈极显著相关,说明这些性状间存在共同遗传基础。
2. 5 蒸煮食味品质性状的QTL定位结果
如表3和圖1所示,在第6染色体共检测出3个蒸煮食味品质性状的主效QTL,包括直链淀粉含量QTL(qAC-6-1)、胶稠度QTL(qGC-6-1)和碱消值QTL(qGT-6-1)各1个,其中,qAC-6-1位于第6染色体RM589和RM19391之间,加性效应为-4.19,表型变异贡献率为65.8%,增效等位基因来自珍汕97B;qGC-6-1也位于第6染色体RM589和RM19391之间,加性效应为12.91,表型变异贡献率为39.3%,增效等位基因来自柳沙油占202;qGT-6-1位于第6染色体RM276和RM7311之间,加性效应为1.78,表型贡献率为44.3%,增效等位基因来自柳沙油占202。综上所述,直链淀粉含量和胶稠度的QTL均位于第6染色体RM589和RM19391之间,且碱消值的QTL定位结果与出糙率和精米率的QTL定位结果相同,均位于第6染色体RM276和RM7311之间。结合上述相关性分析结果,直链淀粉含量与胶稠度呈极显著负相关;出糙率、精米率和碱消值均呈极显著正相关,说明前2个性状和后3个性状分别共享相同或相似的分子遗传机制,以支撑表型的高度相关性。
3 讨论
稻米品质是一个综合性状,其中碾磨加工品质性状对稻米品质影响较大,直接影响到稻米市场价格和销售(郭桂英等,2017)。本研究在第6染色体检测出1个出糙率QTL qPBR-6-1,其位于RM276和RM7311标记之间。周勇等(2013)利用染色体单片段代换系将糙米率QTL qBRR6-1定位在RM8258和S6-7区段。由于该区段与RM276~RM7311标记区域重叠,说明本研究结果与周勇等(2013)的研究结果相似。方雅洁等(2018)在第6染色体检测出1个糙米率QTL qBBR6,其所在位置与已克隆的粒型基因GS6(Sun et al.,2013)一致。梅德勇等(2012)在第6染色体检测出1个糙米率QTL qBRR6,其位于RM190和RM587标记之间。本研究检测出的出糙率qPBR-6-1位于2个qBBR6的下游,其是否为新的出糙率QTL仍需做进一步鉴定。本研究检测出的精米率QTL qMRR-6-1与方雅洁等(2018)检测出的精米率QTL qMRR6.2位置(Chr.6:3490962~4782482)較接近,且该位置附近存在淀粉合成基因SSG6(Matsushima et al.,2016),本研究相关性分析结果也显示,精米率与直链淀粉含量呈极显著负相关;另一个精米率主效QTL qMRR-6-2与方雅洁等(2018)检测出的精米率QTL qMRR6.1(Chr.6:852588~1508680)所在位置存在重叠。
外观品质性状是评价稻米品质优劣的重要标准。本研究发现,粒宽QTL qGW-5-1和长宽比QTL qRLW-5-1均位于第5染色体RM169和RM574标记之间。结合前人研究报道(Weng et al.,2008)发现,该区域包含粒宽基因GW5。本研究检测出的垩白率主效QTL qCGR-5-1和垩白度主效QTL qC-5-1与Liu等(2012)检测出的垩白率主效QTL qJPGC-5位置(Chr.5:RM289~RM3437)非常接近;垩白粒率主效QTL qCGR-6-1与晁园等(2012)在第6染色体检测出的垩白率QTLqCR6-4位置(Chr.6:RM19382~RM510)接近。
随着人民生活水平的提高,提高稻米蒸煮品质逐渐成为水稻优质化育种改良的重要目标(邵高能等,2009)。Wang等(2007)研究表明,米饭的蒸煮食味品质主要受Wx和alk基因调控,其中Wx基因主要调控直链淀粉含量和胶稠度,而alk基因主要调控碱消值和糊化温度。本研究检测出直链淀粉含量主效QTL qAC-6-1和胶稠度主效QTL qGC-6-1均位于第6染色体RM589和RM19391标记之间,碱消值主效QTL qGT-6-1定位于第6染色体RM276和RM7311标记之间。结合晁园等(2012)的研究报道,推测直链淀粉含量主效QTL qAC-6-1和碱消值主效QTL qGT-6-1分别位于Wx和alk基因所在位置。且本研究相关性分析结果与邵高能等(2009)相关性分析结果均显示,直链淀粉含量与胶稠度呈极显著负相关,表明二者共享相同或相似的分子遗传机制,以支撑表型的高度相关性。
大量研究表明,利用重组自交系检测品质相关性状的QTL时,存在检测所得目标QTL较少的问题。如邵高能等(2009)利用重组自交系对直链淀粉含量、糊化温度及胶稠度等性状进行QTL定位,结果仅检测到6个具有显著加性效应的QTL;张上都等(2016)利用重组自交系对糙米率和整精米率进行QTL定位,结果仅检测出2个相关的QTL。本研究也发现类似问题,利用重组自交系对12个稻米品质性状进行QTL定位,结果仅检测到11个QTL,推测其主要原因是柳沙油占202和珍汕97B两亲本均为籼稻,亲缘关系较近,二者间SSR标记的多态性较低(张启军,2004)。本研究从496个SSR标记中筛选获得56个在两亲本中具有较好多态性的标记,多态性频率仅为11.29%。亲本间亲缘关系较近,能用于构建遗传连锁图的多态性标记较少,遗传连锁图不够细密,因此,只能检测到少数具有较大遗传效应的QTL。在今后的研究中,应加强对亲缘关系较远亲本材料的发掘利用,以获得更多多态性标记用于构建更细密的遗传连锁图,提高品质性状的QTL检测效率。
4 结论
重组自交系群体12个稻米品质性状存在互作效应,与QTL定位结果相互印证。鉴定所得主效QTL可用于水稻稻米品质相关性状的遗传改良。
参考文献:
蔡薇,陈彦成,黄丽华,支添添,任春梅. 2017. 水稻HGO基因启动子的克隆及转化鉴定[J]. 江西农业学报,29(3):18-21. [Cai W,Chen Y C,Huang L H,Zhi T T,Ren C M. 2017. Cloning,transformation and identification of gene HGO promoter in rice[J]. Acta Agriculturae Jiangxi,29(3):18-21.]
晁园,冯付春,高冠军,朱雪萍,何予卿. 2012. 利用重组自交系群体定位水稻品质相关性状的QTL[J]. 华中农业大学学报,31(4):397-403. [Chao Y,Feng F C,Gao G J,Zhu X P,He Y Q. 2012. Mapping QTLs related with rice qualities of appearance,cooking and eating using a recombinant inbred line population[J]. Journal of Huazhong Agricultural University,31(4):397-403.]
方先文,张云辉,肖西林,张所兵,林静,汪迎节. 2017. 基于重组自交系群体的水稻粒形QTL定位[J]. 江苏农业学报,33(2):241-247. [Fang X W,Zhang Y H,Xiao X L,Zhang S B,Lin J,Wang Y J. 2017. Mapping of QTLs for grain shape using recombinant inbred lines in rice(Oryza sativa L.)[J]. Jiangsu Journal of Agricultural Sciences,33(2):241-247.]
方雅洁,朱亚军,吴志超,陈凯,申聪聪,石英尧,徐建龙. 2018. 全基因组关联定位籼稻种质资源外观和加工品质QTL[J]. 作物学报,44(1):32-42. [Fang Y J,Zhu Y J,Wu Z C,Chen K,Shen C C,Shi Y Y,Xu J L. 2018. Genome-wide association study of grain appearance and milling quality in a worldwide collection of Indica rice germplasm[J]. Acta Agronomica Sinica,44(1):32-42.]
高维维,陈思平,王丽平,陈立凯,郭涛,王慧,陈志强. 2017. 稻米蒸煮品质性状与分子标记关联研究[J]. 中国农业科学,50(4):599-611. [Gao W W,Chen S P,Wang L P,Chen L K,Guo T,Wang H,Chen Z Q. 2017. Association analysis of rice cooking quality traits with molecular mar-kers[J]. Scientia Agricultura Sinica,50(4):599-611.]
高振宇,曾大力,崔霞,周奕華,颜美仙,黄大年,李家洋,钱前. 2003. 水稻稻米糊化温度控制基因Alk的图位克隆及其序列分析[J]. 中国科学(C辑),33(6):481-487. [Gao Z Y,Zeng D L,Cui X,Zhou Y H,Yan M X,Huang D N,Li J Y,Qian Q. 2003. Graphic cloning and sequence analysis on the control gene Alk of rice paste temperature[J]. Chinese Science(Series C),33(6):481-487.]
郭桂英,王青林,马汉云,鲁伟林,沈光辉,全瑞兰,余新春,扶定,霍二伟. 2017. 碾磨品质对籼稻食味品质的影响[J]. 天津农业科学,23(6):40-44. [Guo G Y,Wang Q L,Ma H Y,Lu W L,Shen G H,Quan R L,Yu X C,Fu D,Huo E W. 2017. Effect of milling quality on eating quality of indica rice[J]. Tianjin Agricultural Sciences,23(6):40-44.]
胡霞,石瑜敏,贾倩,徐琴,王韵,陈凯,孙勇,朱苓华,徐建龙,黎志康. 2011. 影响水稻穗部性状及籽粒碾磨品质的QTL及其环境互作分析[J]. 作物学报,37(7):1175-1185. [Hu X,Shi Y M,Jia Q,Xu Q,Wang Y,Chen K,Sun Y,Zhu L H,Xu J L,Li Z K. 2011. Analyses of QTLs for rice panicle and milling quality traits and their interaction with environment[J]. Acta Agronomica Sinica,37(7):1175-1185.]
黄发松,孙宗修,胡培松,唐绍清. 1998. 食用稻米品质形成研究的现状与展望[J]. 中国水稻科学,12(3):172-176. [Huang F S,Sun Z X,Hu P S,Tang S Q. 1998. Present situations and prospects for the research on rice grain quality forming[J]. Chinese Journal of Rice Science,12(3):172-176.]
刘家富,奎丽梅,朱作峰,谭禄宾,王桂娟,黎其万,束继红,孙传清. 2007. 普通野生稻稻米加工品质和外观品质的QTL定位[J]. 农业生物技术学报,15(1):90-96. [Liu J F,Kui L M,Zhu Z F,Tan L B,Wang G J,Li Q W,Shu J H,Sun C Q. 2007. Identification of QTLs associated with processing quality andappearance quality of common wild rice(Oryza rufipogon Griff.)[J]. Journal of Agricultural Biotechnology,15(1):90-96.]
梅德勇,朱玉君,樊叶杨. 2012. 籼稻稻米碾磨与外观品质性状的QTL定位[J]. 遗传,34(12):1591-1598. [Mei D Y,Zhu Y J,Fan Y Y. 2012. Mapping QTL for rice milling and appearance quality traits in indica rice[J]. Hereditas,34(12):1591-1598.]
梅捍卫,罗利军,郭龙彪,王一平,余新桥,应存山,黎志康. 2002. 水稻加工品质数量性状基因座(QTIs)分子定位研究[J]. 遗传学报,29(9):791-797. [Mei H W,Luo L J,Guo L B,Wang Y P,Yu X Q,Ying C S,Li Z K. 2002. Molecular mapping of QTLs for rice milling yield traits[J]. Acta Genetica Sinica,29(9):791-797.]
穆平,郭詠梅,刘家富,卢义宣,李自超. 2007. 稻米外观和碾磨品质QTL定位及其与土壤水分环境互作分析[J]. 农业生物技术学报,15(4):654-660. [Mu P,Guo Y M,Liu J F,Lu Y X,Li Z C. 2007. QTL mapping and QTL environments interactions of grain milling and appearance quality traits in rice under upland and lowland environments[J]. Journal of Agricultural Biotechnology,15(4):654-660.]
彭强,张大双,吴健强,张玉珊,王际凤,黄培英,朱速松. 2016. 水稻粒形性状QTL定位分析[J]. 西南农业学报,29(8):1766-1770. [Peng Q,Zhang D S,Wu J Q,Zhang Y S,Wang J F,Huang P Y,Zhu S S. 2016. QTL analysis of grain shape trait of rice[J]. Southwest China Journal of Agricultural Sciences,29(8):1766-1770.]
邵高能,唐绍清,焦桂爱,罗炬,唐傲,胡培松. 2009. 稻米蒸煮品质性状的QTL定位[J]. 中国水稻科学,23(1):94-98. [Shao G N,Tang S Q,Jiao G A,Luo J,Tang A,Hu P S. 2009. Mapping of QTL for cooking quality traits of rice[J]. Chinese Journal Rice Science,23(1):94-98.]
张昌泉,赵冬生,李钱峰,顾铭洪,刘巧泉. 2016. 稻米品质性状基因的克隆与功能研究进展[J]. 中国农业科学,49(22):4267-4283. [Zhang C Q,Zhao D S,Li Q F,Gu M H,Liu Q Q. 2016. Progresses in research on cloning and functional analysis of key genes involving in rice grain quality[J]. Scientia Agricultura Sinica,49(22):4267-4283.]
张启军. 2004. 利用SSR和四个测序水稻品种构建分子图谱及QTLs分析[D]. 雅安:四川农业大学. [Zhang Q J. 2004. Construction of SSR linkage maps between four sequenced rice varieties and mapping of quantitative trait loci[D]. Ya’an:Sichuan Agricultural University.]
张上都,彭强,张大双,吴健强,杨林,朱速松. 2016. 利用RIL群体进行稻米加工品质QTL定位分析[J]. 贵州农业科学,44(6):8-10. [Zhang S D,Peng Q,Zhang D S,Wu J Q,Yang L,Zhu S S. 2016. QTL analysis for milling quali-ty of rice by using RIL population[J]. Guizhou Agricultural Sciences,44(6):8-10.]
张月雄,梁海福,秦钢,马增凤,岑贞陆,刘驰,罗同平,韦敏益,李振经,李容柏,黄大辉. 2018. 籼稻品种9311抗白叶枯基因鉴定和定位[J]. 分子植物育种,16(2):460-465.[Zhang Y X,Liang H F,Qin G,Ma Z F,Cen Z L,Liu C,Luo T P,Wei M Y,Li Z J,Li R B,Huang D H. 2018.Identification and mapping of a bacterial blight resistance gene in Indica CV.9311[J]. Molecular Plant Breeding,16(2):460-465.]
周勇,高云,朱松松,朱金燕,王军,裔传灯,仲维功,梁国华. 2013. 基于染色体单片段代换系的水稻糙米率QTL的定位[J]. 华北农学报,28(6):1-5. [Zhou Y,Gao Y,Zhu S S,Zhu J Y,Wang J,Yi C D,Zhong W G,Liang G H. 2013. QTL mapping of brown rice rate using chromosome single segment substituted lines in rice[J]. Acta Agriculturae Boreali-Sinica,28(6):1-5.]
Guo T,Liu X L,Wan X Y,Weng J F,Liu S J,Liu X,Chen M J,Li J J,Su N,Wu F Q,Cheng Z J,Guo X P,Lei C L,Wang J L,Jiang L,Wan J M. 2011. Identification of a stable quantitative trait locus for percentage grains with white chalkiness in rice(Oryza sativa)[J]. Journal of Integrative Plant Biology,53(8):598-607.
Li Y B,Fan C C,Xing Y Z,Jiang Y H,Luo L J,Sun L,Shao D,Xu C J,Li X H,Xiao J H,He Y Q,Zhang Q F. 2011. Natural variation in GS5 plays an important role in regulating grain size andyield in rice[J]. Nature Genetics,43(12):1266-1269.
Liu X,Wang Y,Wang S W. 2012. QTL analysis of percen-tage of grains with chalkiness in Japonica rice(Oryza sativa)[J]. Genetics and Molecular Research,11(1):717-724.
Matsushima R,Maekawa M,Kusano M,Tomita K,Kondo H,Nishimura H,Crofts N,Fujita N,Sakamoto W. 2016. A-myloplast membrane protein SUBSTANDARD STARCH GRAIN6 controls starch grain size in rice endosperm[J]. Plant Physiology,170(3):1445-1459.
Song X J,Huang W,Shi M,Zhu M Z,Lin H X. 2007. A QTL forrice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase[J]. Nature Genetics,39(5):623-630.
Sun L J,Li X J,Fu Y C,Zhu Z G,Tan L B,Liu F X,Sun X Y,Sun X W,Sun C Q. 2013. GS6,a member of the GRAS gene family,negatively regulates grain size in rice[J]. Journal of Integrative Plant Biology,55(10):938-949.
Wang L Q,Liu W J,Xu Y,He Y Q ,Luo L J,Xing Y Z,Xu C G,Zhang Q F. 2007. Genetic basis of 17 traits and viscosity parameters characterizing the eating and cooking quality of rice grain[J]. Theoretical Applied Genetics,115(4):463-476.
Wang S K,Lis,Liu Q,Wu K,Zhang J,Wang S S,Wang Y,Chen X B,Zhang Y,Gao C X,Wang G F,Huang H X,Fu X D. 2015. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality[J]. Nature Genetics,47(8):949-954.
Wang Z Y,Zheng F Q,Shen G Z,Gao J P,Snustad D P,Li M G,Zhang J L,Hong M M. 1995. The amylose content in rice endosperm is related to the post-transcriptional regulation of the waxy gene[J]. The Plant Journal,7(4):613-622.
Weng J F,Gu S H,Wan X Y,Gao H,Gut T,Su N,Lei C L,Zhang X,Cheng Z J,Guo X P,Wang J L,Jiang L,Zhai H Q,Wan J M. 2008. Isolation and initial characterization of GW5,a major QTL associated with rice grain width and weight[J]. Cell Research,18(12):1199-1209.
Zhou L J,Chen L M,Jiang L,Zhang W W,Liu L L,Liu X,Zhao Z G,Liu S J,Zhang L J,Wang J K,Wan J M. 2009. Finemapping of the grain chalkiness QTL qPGWC-7 in rice(Oryza sativa L.)[J]. Theoretical and Applied Gene-tics,118(3):581-590.
(責任编辑 陈 燕)
