紫金山针阔混交林群落特征
2019-09-10陈霞袁在翔金雪梅关庆伟朱军戴康龙赵宸
陈霞 袁在翔 金雪梅 关庆伟 朱军 戴康龙 赵宸



摘要:了解紫金山针阔混交林物种组成、优势种、植物区系、生活型、多样性等群落特征,可为紫金山森林资源的监测与管理提供一定参考价值。文章基于针阔混交林lhm2典型样地的植被调查数据,运用数量生态学方法,对上述群落特征进行研究。结果表明:1)针阔混交林群落维管束植物共计50科89属105种,其中蕨类植物2科2种2属,种子植物48科87属103种;2)群落优势种为马尾松、栓皮栎,但二者的林下更新较差;3)植物属的区系成分以温带分布为主,其次为热带分布;4)生活型以高位芽植物为主,其他生活型所占比例相对较少;5)群落乔木层与乔木更新层的物种多样性指数较为相似,并且显著高于其他层次。
关键词:物种多样性,重要值,植物区系,生活型,针阔混交林
DOI: 10.12169/zgcsly.2019.05.23.0003
紫金山针阔混交林是在人为干扰相对较弱的 阔叶树种入侵马尾松纯林而形成的,在我国北亚情况下,经过约65年的演替,由栓皮栎、朴树等 热带地区具有一定的代表性[1-2]。近年来对紫金山针阔混交林的研究多集中在演替动态[1]、空间结构[3-4]、植物多样性[4-5]、土壤性质[6-7]等方面,对群落中物种组成、植物区系、生活型等特征的研究较少。因此,笔者基于紫金山针阔混交林1hm2典型样地的植被调查数据,运用数量生态学方法,对针阔混交林物种组成、优势种、区系、生活型、多样性等群落特征进行研究,旨在更好地了解森林群落现状,为紫金山森林资源监测与管理提供一定的参考价值。
1研究地概况
紫金山(118°48'24”-118°53 704”E.32°01'57”-32°06 ’15”N)位于南京市玄武区,主峰海拔高度为448.9 m,相对高度为420 m,总面积为2 970 hm2,属于北亚热带季风氣候,年降水量为1 000~1 050 mm,年均气温15.4℃。土壤类型以黄棕壤和灰棕壤为主,酸性至微酸性。林分类型有落叶阔叶林、马尾松纯林和针阔混交林。针阔混交林主要乔木为马尾松(Pinus ma.ssoniana)、栓皮栎(Quercus variadilis)、朴树(Celtissinensts)、紫薇(Lagerstroemia indica);主要灌木为山莓(Rubus corchorifolius]、菝禊(Smilax chi-na)、六月雪(Serissa japonica)、雀梅藤(Sageretia thea);主要草本为苔草(Carex chinen-sis)、麦冬(Liriope graminifolia)、狭叶鳞毛蕨(Dryopteris angustifrons)、野青茅(Deyeuxiaarundinacea)。
2研究方法
2.1样地设置与调查方法
于2014年10月,在紫金山南麓的针阔混交林中用全站仪和罗盘仪划定典型样地1hm2.样地内GPS测定点的位置为118050'23.7”E.32004'03.9”N,海拔230 m,上坡位。运用相邻格子法把典型样地划分25个20 mx20 m的调查样方。在各调查样方内进行每木调查,并对样地内胸径≥5cm的树木分别定位,记录所研究林分中每株个体的位置,以坐标值表示,分别记录每株林木的树种、胸径(≥5 cm)、树高、冠幅和坐标定位,坐标值用距离(m)直接表示。在每个20 m×20 m调查样方内随机设置20个2 mx2 m的小样方,调查并记录每个小样方内更新幼树、灌木、草本和藤本植物的种名、基径、平均高度、盖度等指标。
2.2群落特征值计算方法
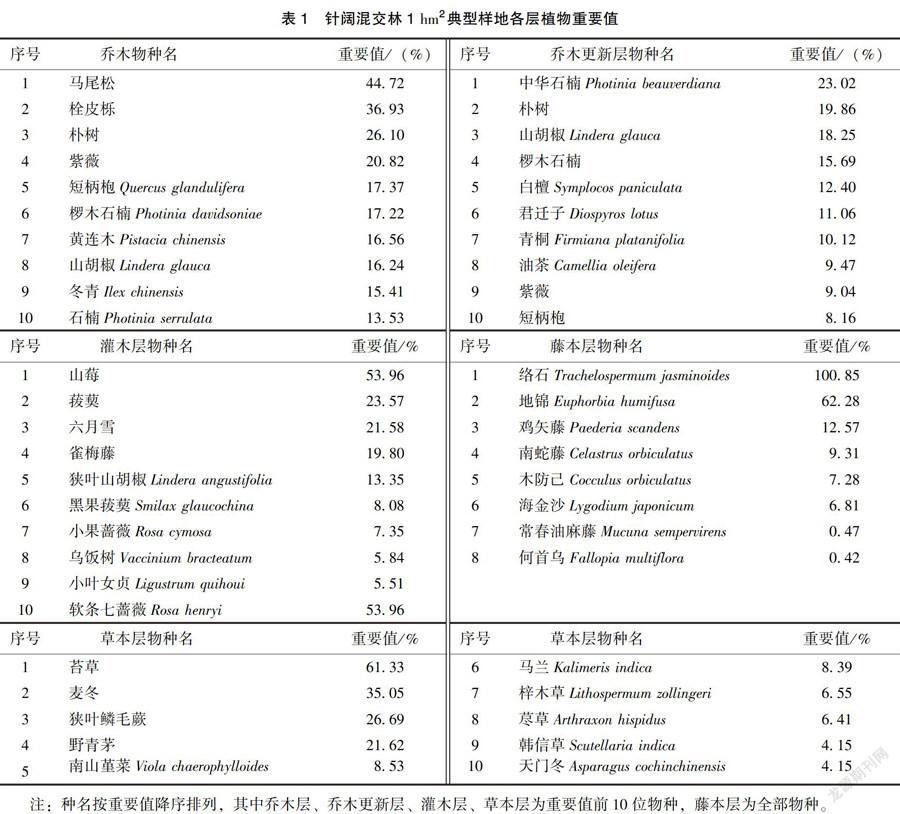
1)植物重要值测定。通过重要值大小描述乔木层、乔木更新层、灌木层、藤本层、草本层的植物种群优势度,进而确定各层主要植物种群。各层植物重要值的计算公式参考梁士楚等[8]的《生态学》,其结果见表1。
2)植物区系分析。维管束植物中种子植物属的分布区类型划分参照吴征镒[9]的《中国种子植物属的分布区类型》文献中的划分标准,蕨类植物属的分布区类型划分则参考《中国植物志》(电子版网站:http://frps.iplant.cn/)的划分标准。
3)植物生活型分析。应用Raunkier[10]提出的植物生活型分类系统进行植物生活型的统计,并计算各植物生活型的百分率,汇编成全国划分标准统一的植物生活型谱。
4)物种多样性测定。物种多样性选用Patrick丰富度指数(R)、Simpson多样性指数(D)、Shannon-wiener多样性指数(H’)、Pielou均匀度指数(E)进行表征,具体计算公式见马克平[11]等《北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数》。
本研究用Excel 2016和R 3.4.1统计分析软件进行数据处理与分析。
3结果与分析
3.1植物区系特征分析
3.1.1种类组成
通过野外调查和数据整理,紫金山针阔混交林典型样地内共出现维管束植物105种,隶属50科89属,分别占紫金山植物总科数的30. 12%,总属数的14. 93%,总种数的7.82%(表2)。含10种以上的科有蔷薇科(Rosaceae,13种);含5~10种的科有豆科(Leguminosae,7种)、壳斗科(Fagaceae,6种)、百合科(Liliaceae,5种);含2~4种的科有木犀科(Oleaceae,4种)、大戟科(Euphorbiaceae.4种)等17个科,共含45个物种;单种科有松科( Pinaceae)、柏科(Cupres-saceae)、金缕梅科(Hamamelidaceae)等29个科。含物种数最多的属有栎属( Quercus)、石楠属(Photinia)、蔷薇属(Rosa)、堇菜属(Viola),各含4个物种;其次为冬青属(Ilex)、女贞属(Ligustrum)、山胡椒属(Lindera)、樱属(Cera.sus)、菝葜属(Smilax)、悬钩子属( Rubus)、胡颓子属(Elaeagnus)、大戟属(Eu-phorbia),各含2个物种;单种属有松属( Pinus)、圆柏属(Sabina)等77个属,占总属数的86. 52%。此外,乔木层有38个物种,其中优势种为马尾松、栓皮栎,主要种有朴树、紫薇等:乔木更新层有37个物种,重要值前4位的物种是中华石楠、朴树、山胡椒、椤木石楠;灌木层有31个物种,主要种为山莓、菝葜、六月雪、雀梅藤等;藤本层有8个物种,其中以络石、地锦较为常见:草本层有22个物种,主要种为苔草、麦冬、狭叶鳞毛蕨、野青茅等。
3.1.2区系成分
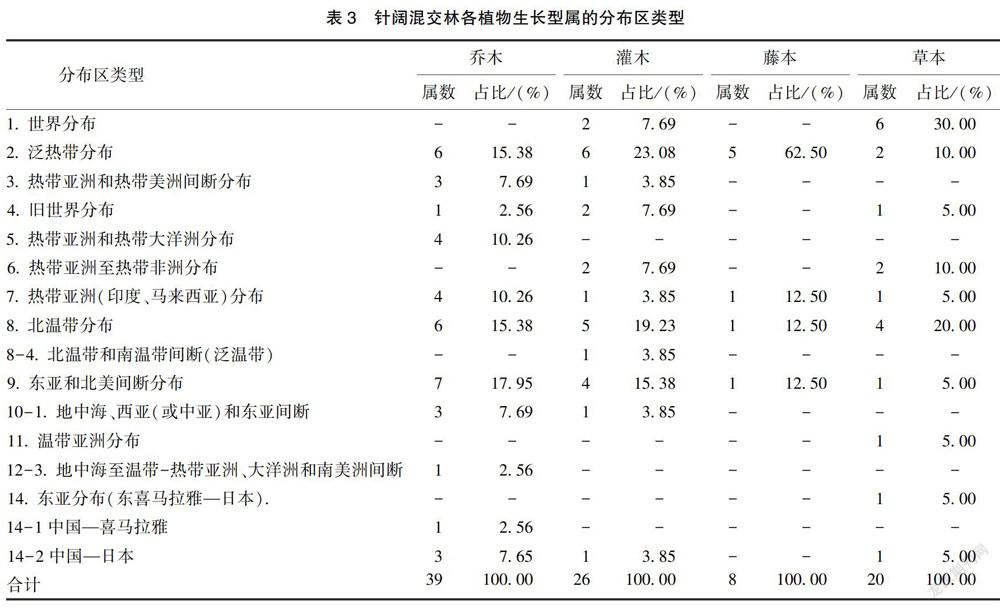
在植物区系研究中,属的分布区类型能够比较具体反映植物界在进化过程中的分异情况和地理特征[13]。根据植物属的分布区类型划分标准,针阔混交林典型样地89属维管束植物共划分11个分布区类型和5个变型。除世界分布类型外,属的地理分布以温带分布为主(8~ 14型),共计43属,占48. 31%;热带分布(2~7型)次之,共有38属,占42. 70%;世界分布仅有8属,占8. 99%。其中,热带分布属中以泛热带分布所占数量最多,共计18属,占维管束植物总属数的20. 22%:而温带分布属中则以北温带分布及其变型所占比例最多,为维管束植物总属数的17. 98%。由表3可知,乔木和草本的属分布区类型均以温带分布为主,灌木的属分布区类型中热带分布与温带分布所占比例相同,藤本的属分布区类型则以热带分布为主。
3.2生活型分析
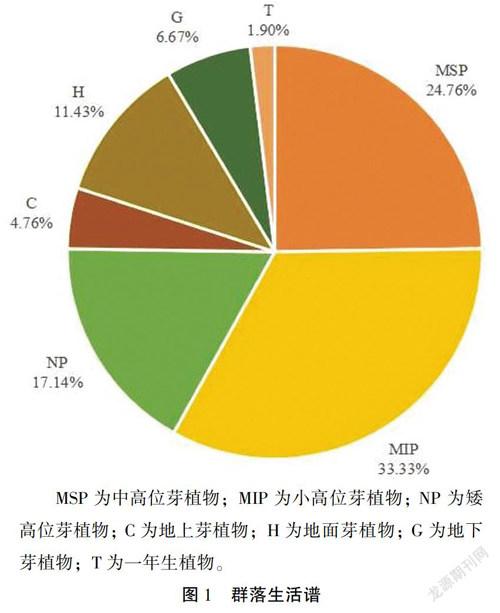
对紫金山针阔混交林样地调查的105种植物的生活型进行统计分析,结果如图1所示。在105种维管束植物中,高位芽植物种类最多(79种),占总物种数的75. 24%;地面芽植物次之(12种),占11. 43%;地下芽和地上芽植物较少(7种和5种),占比分别为6. 67%和4.76%; 一年生植物最少,仅占总物种数的1. 90%。在高位芽植物中,以小高位芽(35种)和中高位芽(26种)植物较多,矮高位芽(18种)植物次之,大高位芽(0种)植物缺乏。
将本研究区植物生活型谱与郭泉水[14]的《中国主要森林群落植物生活型谱的数量分类及空间分布格局的研究》文中的亚热带常绿阔叶林(Ⅱ)、北亚热带常绿、落叶阔叶林(Ⅲ)和暖温带落叶栎林(Ⅳ)的植物生活型譜比较(表4)。表4数据显示,本研究区的植物生活型谱更偏向于北亚热带常绿、落叶阔叶林(Ⅲ)生活型谱特征,即以高位芽植物为主,含有一定比例的地面芽和地下芽植物,同时一年生植物极少。
3.3物种多样性
物种多样性是群落中物种丰富度和分布均匀性的综合反映,体现了群落结构类型、组织水平、发展阶段、稳定程度和生境差异[15]。对针阔混交林乔木层、乔木更新层、灌木层、藤本层、草本层的物种多样性进行分析,结果见表5。各层次丰富度指数表现为:乔木层>乔木更新层>灌木层>草本层>藤本层;而多样性指数、均匀度指数则表现为:乔木更新层>乔木层>灌木层>草本层>藤本层。
4结论与讨论
4.1针阔混交林植被特征
紫金山针阔混交林群落内维管束植物共计50科89属105种,其中蕨类植物2科2种2属,种子植物48科87属103种。从生长型角度看,乔木25科39属44种,灌木18科26属31种,藤本8科8属8种,草本15科20属22种。群落优势种为马尾松、栓皮栎,但在乔木更新层的1046株幼树幼苗中,马尾松、栓皮栎分别仅占0%、1. 72%,表明在当前演替阶段马尾松、栓皮栎的幼树幼苗更新较为困难,这与袁在翔[16]等研究结果相似,其主要原因是马尾松和栓皮栎均为阳性树种,而林冠郁闭度较大,导致光线较差,抑制了种子萌发和幼树幼苗生长。
针阔混交林植物区系的地理成分较为复杂,属的地理分布类型中热带成分与温带成分的比值( R/T)为0.88,表现出显著的热带向温带过渡的特征,符合研究区所处的北亚热带地理位置,这与安树青[17]等、王春[12]等研究结果相一致。植物生活型谱以高位芽植物为主,其次为地面芽植物,含有一定比例的地下芽和地上芽植物,一年生植物种类极少。在高位芽植物中,大高位芽植物缺乏,并且常绿成分与落叶成分的比值为0. 47,基本符合中国北亚热带次生森林植被的特征[18],其原因可能与本区域水热条件不足和人类活动的影响相关[19]。
此外,群落中乔木层与乔木更新层物种多样性指数较为接近,且均显著高于其他层次,这与朱军[4]等、吴迪[5]等研究结果存在一定的差异。前者是因为更新层的幼树幼苗大多为实生苗,其物种类别与分布和乔木层的母树关系密切,而物种多样性指数是物种丰富度和均匀度的函数[11],所以乔木层与更新层的物种多样性指数较为相似:后者是因为本研究把幼树幼苗单独归类为乔木更新层,而不是纳入灌木层,造成灌木层的总物种数和分布均匀度存在一定程度的下降,故而区别于以往灌木层物种多样性指数最大的特征。
4.2从群落演替角度分析针阔混交林植被特征
当前演替阶段针阔混交林的植被特征符合安树青[20]等研究得出的紫金山次生森林群落的演替趋势,即马尾松群落一马尾松、落叶阔叶树群落一落叶阔叶树、马尾松群落一落叶阔叶树群落,主要体现为3个方面。
1)马尾松种群进一步衰退,林下更新为零。1951年马尾松种群个体数量[21]为1700株/hm2,其中苗木为300株/hm2; 1981年马尾松种群个体数量[22]约2 318株/hm2,其中苗木为500株/hm2;2002年马尾松种群个体数量[1]约309株/hm2.其中苗木为0株/hm2; 2010年马尾松种群个体数量[23]约379株/hm2; 2014年马尾松种群个体数量[1]约127株/hm2,其中苗木株数为0。
2)阔叶树种的个体数量所占比例总体上呈增加趋势。1951年、1981年、2002年、2010年、2014年马尾松群落阔叶树种个体数所占比例分别为82. 4%[21], 83. 8%[22], 90. 4%[1,96. 5%[23],95. 04%。
3)多样性指数呈现波动性变化特征,具有一定的增加趋势[24]。1951年、1981年、2002年、2010年、2014年木本植物Simpson多样性指数分别为0. 936[1],0.799[1],0. 927[1],0.914[23],0.972; Shannon - wiener多样性指数分别为1. 381[1],0.985[1],1.346[1],2.923[23],3. 953。
参考文献
[1]徐驰,刘茂松,张明娟,等.南京灵谷寺森林50年来的动态变化研究[J].植物生态学报,2004,28(5):601-608.
[2]陈霞,袁在翔,金雪梅,等.紫金山针阔混交林主要树种空间分布格局及种间关联性[J].南京林业大学学报(自然科学版),2018,42(6):84-90.
[3]湛斌,闫新利,陈斌,等.紫金山针阔混交林空间结构特征分析[J]西北林学院学报,2016,31(5):224-229.
[4]朱军,湛斌,陈霞,等紫金山针阔混交林空间结构特征与植物多样性分析[J].中南林业科技大学学报,2018,38(1):75 -80,93.
[5]吴迪,湛斌,闫新利,等.紫金山栓皮栎枫香混交林群落结构与物种多样性[J].安徽农业大学学报,2017,44(1):50-54.
[6]姜爱国,万福绪,胡菲.南京紫金山灵谷寺不同林地土壤抗蚀性研究[J].水土保持研究,2018,25(1):12-16.
[7]袁在翔南京紫金山2种典型林分土壤碳库与养分特征[D]南京:南京林业大学.2017.
[8]梁士楚,李铭红.生态学[M].武漢:华中科技大学出版社.2015.
[9]吴征镒中国种子植物属的分布区类型[J].云南植物研究,1991,(增4):1-139.
[10] RAUNKIAER G.The life Forms of Plants and Statistical PlantGeography[M].Oxford: Clarendon press, 1934.
[11]马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数[J].生态学报,1995(3):268-277.
[12]王春.南京紫金山植物区系与植物资源研究[D].南京:南京林业大学.2009
[13]吴征镒,孙航,周浙昆,等.中国种子植物区系地理[J].生物多样性,2011,19(1):124-124.
[14]郭泉水,江洪,王兵,等.中国主要森林群落植物生活型谱的数量分类及空间分布格局的研究[J].生态学报,1999,19(4):573-577.
[15]张全国,张大勇生物多样性与生态系统功能:最新的进展与动向[J].生物多样性,2003,11(5):351-363.
[16]袁在翔,金雪梅,马婷瑶,等.南京灵谷寺栓皮栎种群结构与动态[J].生态学杂志,2017,36(6):1488-1494.
[17]安树青,赵儒林紫金山次生森林植被特征分析[J].植物生态学报,1990,14(1):13 -22.
[18]安树青,赵儒林.中国北亚热带次生森林植被的特征分析[J].南京大学学报(自然科学),1991,27(2):323-331
[19]高贤明,陈灵芝植物生活型分类系统的修订及中国暖温带森林植物生活型谱分析[J].植物学报,1998,40(6):553-559.
[20]安树青,王峥峰,刘志礼土壤因子对次生森林群落演替的影响[J].生态学报,1997(1):45-50
[21]曲仲湘,文振旺,朱克贵.南京灵谷寺森林现况的分析[J].植物学报,1952(1):18-49
[22]熊文愈,韩福庆,姚琢.南京灵谷寺森林的变化分析[J].南京林产工业学院学报,1983,7(2):1-23.
[23]曹静.南京紫金山森林植物多样性及其价值评估[D].南京:南京林业大学.2011.
[24]陈霞,湛斌,朱军,等.紫金山马尾松枫香混交林多样性研究[J].中国城市林业,2018,16(1):64-68.
收稿日期:2019-05-23
基金项目:2015年南京市建设系统科研计划项目“紫金山典型森林群落的植物多样性大样地长期观测研究”(Ks1501)
第一作者:陈霞(1975 -),女,硕士,高级工程师,研究方向为森林管理。E-mail: 157168037@ qq.com
通信作者:关庆伟(1964-),男,教授,博士生导师,研究方向为森林生态与经营。E-mail:guanjapan999@ 163.com
