狭域特有植物元江素馨的种群结构及动态特征
2019-09-10李娟邹琳吕亚媚肖明昆杜凡
李娟 邹琳 吕亚媚 肖明昆 杜凡


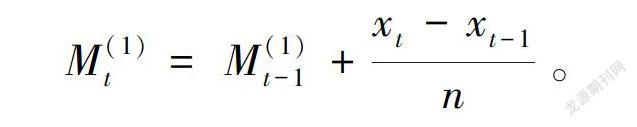

摘要: 元江素馨(Jasminum yuanjiangense)为云南省元江河谷狭限分布的特有植物,该研究采用典型选样法在元江素馨较为集中分布地布置4个样方,包含干热灌丛、稀树灌木草丛2个植被类型,调查分析种群年龄结构及存活曲线,量化种群结构类型,比较各局域元江素馨种群的差异性,并运用时间序列预测种群数量动态,以揭示元江素馨种群结构及动态特征。结果表明:(1)元江素馨种群属于增长型,Vpi′=0.004 1,表明该种群趋近于稳定型;种群结构在发展过程中存在一定的波动性,种群生长前期个体数多于生长后期;存活曲线趋于Deevey-Ⅲ型和Deevey-Ⅰ型;幼苗充足,死亡率高,幼龄阶段到中龄阶段个体生长发育受阻。(2)时间序列预测分析表明,元江素馨种群具备一定的恢复能力,未来2、4个龄级时间后种群个体数均呈现小幅度的增加趋势。(3)各局域元江素馨种群结构存在差异,但依据其年龄结构大致可划分为两类,即样地1、样地2和样地4种群为增长型,存活曲线表现出Deevey-Ⅲ型;样地3的种群为稳定型,存活曲线呈现出Deevey-Ⅰ型,稳定型种群生境较增长型种群生境更适宜于元江素馨的生长。
关键词: 元江素馨, 种群, 静态生命表, 存活曲线, 时间序列预测
中图分类号: Q948文献标识码: A文章编号: 1000-3142(2019)09-1271-10
Abstract: Jasminum yuanjiangense is an narrowly distributed endemic species in Yuanjiang River Valley of Yunnan Province. Four sample plots, belongs to two vegetation types including dry-hot shrubs and savanna were typically selected in the distribution area of J. yuanjiangense. The population age structure and survival curves were created and the type of population structure were analysed by Chen Xiaodes Quantification Method, and predicted population dynamics using time sequence model, and compared the population differences between different habitats. The results were as follows: (1) The population of J. yuanjiangense presented as a growing type and tend to stable(Vpi′=0.004 1). The populations had definite fluctuations during their growth dynamics, and the number of individuals in the pre-growth stage was more than that in the late growth stage. The survival curve of population approached to type Deevey-Ⅲ and Deevey-Ⅰ. The seedlings was abundant, but the mortality was high. The growth and development of individuals were limited from young to middle age. (2) The time sequence model predicted that the population of J. yuanjiangense was able to keep stable and the number of individuals would have a light rising tendency in the coming 2, 4 age classes. (3) The population structure of J. yuanjiangense varies greatly among different sites, but it can be roughly divided into two types according to its age structure. Sample plots 1, 2 and 4 (Habitat 1) displayed growing type with Deevey-Ⅲ survival curve, Plot 3 (Habitat 2) displayed stable type with Deevey-Ⅰ survival curve. The habitat of stable type is more suitable for the growth of J. yuanjiangense than the habitat of growing type.
Key words: Jasminum yuanjiangense, population, static life table, survival curve, time sequence prediction
種群生态学是研究种群数量动态与环境相互作用关系的学科(戈峰,2008)。种群是同一物种占有一定时间和空间的个体集合,是物种的存在单位、繁殖单位和进化单位(薛建辉,2006)。植物种群的年龄结构是指不同年龄组的个体在种群内的比例和配置情况(李博,2000),不仅能反映种群个体数量的多少和分布状况,而且也能反映种群的发生发展动态(韩璐等,2014)。生命表和存活曲线是研究种群结构及动态变化的重要工具(王卓等,2009),可直观反映现有种群各龄级阶段的个体数、死亡数以及种群未来变化趋势(刘国军等,2011)。因此,通过对植物种群结构动态进行研究,了解现有植物种群对环境适应的结果,预测种群未来发生发展趋势,对狭域特有种群的保护和管理具有重要意义(申仕康等,2008)。
元江素馨(Jasminum yuanjiangense)建立于1985年(白佩瑜,1985;中国科学院中国植物志编辑委员会,1992),为木犀科(Oleaceae Hoffmanns & Link)素馨属(Jasminum Dum.)攀援灌木,是云南省元江河谷的土著种,是元江河谷狭域特有种,还是元江国家级自然保护区重要物种监测对象。元江素馨花白色,小而密,是较好的园林绿化植物,但尚未进行开发利用。目前,素馨属植物的研究主要集中于系统学(白佩瑜,1983;刘建林,2005;杨雪等,2006;裴洪光等,2013)、药物化学(张修朋等,2014)及植物生理学(程淑娟等,2012)等方面,但对该属植物种群学、群落学的研究却少见。笔者调查到元江素馨分布于元江干热河谷海拔500~850 m的范围内,该区高温干热,人地矛盾突出,人为干扰严重。在如此恶劣的河谷生境中,影响元江素馨分布、种群的因子有哪些,这些因子如何影响元江素馨种群的发生发展,同时元江素馨种群又是如何适应和响应的。本研究以元江河谷特有物种元江素馨为对象,首先通过分析元江素馨种群年龄结构特征,以及编制静态生命表,绘制种群存活曲线,确定种群结构类型,了解现有种群动态及发展趋势,揭示元江素馨对干热河谷的独特适应性,探讨元江素馨种群的更新和壮大机制;并运用时间序列预测模型分析元江素馨未来种群数量动态,揭示元江素馨种群数量动态变化趋势,为解决自然种群的高死亡率问题和该物种资源的保护与可持续利用提供科学依据。
1材料与方法
1.1 研究区域概况
研究地为云南省中南部元江县,地理坐标为101°39′—102°21′ E, 23°18′—23°54′ N之间,元江素馨分布地土壤属燥红壤,岩石裸露,土壤贫瘠、干燥。研究区位于该区海拔1 000 m以下区域,是我国第一个最具典型特征的干热河谷区域。此区属北热带气候,气温高,年均温为24.1 ℃,极端高温达42.3 ℃,降雨少,蒸发量大,梵风效应强;植被类型以干热河谷灌丛和干热性稀树灌木草丛为主。
1.2 样地设置
根据元江素馨零星或块状分布的特点,样地设置采用典型选样原则,选择元江素馨分布较为集中区域,设置3块20 m × 20 m和1块30 m × 30 m的样地(图1),记录样地的坡位、坡度、坡向等生境因子,每块样地分成5 m×5 m小样方,记录样方内元江素馨的高度、东西冠幅、南北冠幅、株数等因子(表1)。
1.3 种群结构分析
1.3.1 种群年龄结构调查及处理根据元江素馨的生长规律和生活史特点,参考油蒿(Artemisia ordosica)、山麻杆(Alchornea davidii)等(杨洪晓等,2006;曾嘉庆,2016)的分级标准。采用体积立方根计算方法划分元江素馨大小级,即植株东西冠幅(P1)、南北冠幅(P2)和地上部分高度(H)三者乘积的立方根(d),将元江素馨按植株体积的立方根划分为5个大小级,以代替元江素馨的年龄等级。Ⅰ级,0<d≤40 cm,为幼龄阶段;Ⅱ级,40<d≤60 cm;Ⅲ级,60<d≤80 cm,为中龄阶段;Ⅳ级,80<d≤100 cm;Ⅴ级,d>100 cm,为老龄阶段。根据各龄级个体数绘制年龄结构图,判断其种群结构类型。公式为d=3(P1×P2×H)。
1.3.2 静态生命表编制及存活曲线根据野外调查资料,统计各个龄级元江素馨植株数,编制元江素馨静态生命表,根据静态生命表绘制种群存活曲线。本文以龄级和标准化后存活个体数lx为横纵坐标,绘制元江素馨的存活曲线。
1.4 种群结构动态
种群结构动态分析采用陈晓德量化方法(陈晓德,1999),分析元江素馨种群内两个相邻龄级(n,n+1)个体间数量变化动态Vn、元江素馨种群年龄结构动态指数Vpi,在考虑未来外部干扰时,种群龄级数量K及种群各龄级个体数Sn对未来外部干扰存在稀释作用,即K与Sn值越大,其稀释作用越强,故考虑外部干扰时,种群年龄结构动态值为VPi′。
1.5 种群数量动态模型
预测生物种群动态有多种模型。本文采用时间序列一次移动平均法对元江素馨种群年龄结构进行模拟预测(张文辉等,2005;郭其强等,2010),模型如下:
式中,t为龄级,Mt(1)是近期n个观測值在t时刻的平均值,文中t分别取种群经历2、4个龄级,预测元江素馨未来种群个体数量变化趋势。
2结果与分析
2.1 种群年龄结构
2.1.1元江素馨种群数量面积2 100 m2的样地中,记录653株元江素馨,其种群平均密度为每平方米0.311株,各局域种群密度差异明显,以样地2密度最大,达每平方米0.555株,样地3密度最小,仅每平方米0.092株。4块样地平均高依次为样地3(1.07 m)>样地4(0.65 m)>样地1(0.59 m)>样地2(0.50 m)。4块样地中元江素馨最大高度出现于样地1中,为3.34 m(表2)。
2.1.2 元江素馨种群的年龄结构元江素馨某一时间段的种群年龄结构图是对该种群现状客观的反映,同时也可以直观地体现该种群的未来发展变化趋势。将元江素馨各局域种群进行单因素方差分析和LSD多重比较,结果显示差异性极显著,即P=0.003>0.01,各局域元江素馨种群结构差异明显,不同生境对其龄级结构影响较大。由图2可知,除样地3外,各局域元江素馨种群年龄结构均呈金字塔形,表明具有高出生率和低死亡率,种群数量处于增长时期;样地3中年龄结构图呈壶型,幼龄阶段个体数和中龄阶段个体数相差不大,表明种群数量处于稳定状态。
元江素馨种群结构从Ⅰ龄级到Ⅴ龄级个体数依次递减,Ⅰ龄级到Ⅱ龄级个体数波动最大,枯死木多,表明幼苗数量充足,但其很难过渡到Ⅱ龄级;Ⅱ龄级至Ⅳ龄级死亡个体逐渐减少,表明元江素馨个体生长对生存空间及生存资源需求增大,种内及种间竞争使种群死亡率维持在一个较高而相对稳定的水平;Ⅳ龄级和Ⅴ龄级个体数相差较小,表明元江素馨种内、种间竞争基本处于平衡,生存空间及生存资源已经不再是影响种群数量的主要原因。
综上所述,样地1、样地2和样地4中各年龄阶段个体数依次为幼龄阶段(87、153、164)>中龄阶段(48、55、48)>老龄阶段(23、14、24),种群中幼龄阶段个体数多,老龄阶段个体数少,种群处于增长时期;样地3中各年龄阶段个体数为中龄阶段(20)>幼龄阶段(12)>老龄阶段(5),种群中龄阶段个体数较幼龄阶段个体数多,老龄阶段个体数最少,表明种群处于稳定状态。
2.1.3 元江素馨静态生命表及存活曲线元江素馨为天然种群,故采用空间取代时间的方法编制静态生命表。由表3可知,元江素馨种群年龄增加,标准化存活数lx减少。除样地3外,其余样地消失率Kx、死亡率qx随年龄增加而减小,虽出现轻微波动,但总体呈现减小趋势,此趋势同存活曲线一致,即种群生长前期死亡率和消失率大于生长后期,表明种群正处于稳定发展阶段;其种群生命期望ex呈现先增后减的单峰趋势,峰值出现在中龄阶段,即在中龄阶段表现出较高生命期望。样地3中死亡率、消失率呈现先增后减趋势,峰值均出现在Ⅲ龄级阶段,表明此时元江素馨种群内部竞争与外部竞争较强烈,种群内淘汰个体数较多,这与标准化死亡数dx一致。其生命期望值则表现出减小趋势,即在幼龄阶段生命期望值最高。元江素馨种群的死亡率及消失率其最大值出现在Ⅴ龄级,并非因为其死亡率、消失率最大,而是元江素馨在Ⅴ龄级后已达到生理寿命而逐步死亡。
存活曲线是对种群在某一时期内存活状况的反映,是衡量种群增长的基本参数。根据Deevey对存活曲线的划分,有3种类型,即Deevey-Ⅰ型,曲线呈凸型;Deevey-Ⅱ型,曲线呈对角线型;Deevey-Ⅲ型,曲线呈凹型。由图3可知,元江素馨存活曲线表现出Deevey-Ⅲ型(样1、样2、样4)和Deevey-Ⅰ型(样3),Deevey-Ⅲ型表明元江素馨种群幼龄阶段死亡率极高,且只有少数个体能活到生理寿命;Deevey-Ⅰ型表明元江素馨种群在接近生理寿命前仅少量个体死亡,大多数个体基本能达到生理寿命。
2.2 种群结构类型
根据陈晓德量化分析的方法,结果显示, 元江素馨种群相邻龄级个体数量变化动态为V1(72.26%)>V2(45.61%)>V3(41.94%)>V4(16.67%)。数据显示,元江素馨种群各龄级间个体数量变化呈“大—小—大”波动。种群年龄结构的动态指数Vpi为0.611 6,在考虑未来的外部干扰时,种群年龄结构动态与种群年龄级数量及各龄级个体数有关,即Vpi′=0.004 1,接近零,表明种群为增长型种群,但增长速度缓慢。
各局域元江素馨种群相邻个龄级间个体数量存在波动,从表3可以看出,样地种群各龄级增长性大小存在差异,样地1为V1>V3>V2>V4;样地2为V2>V1>V3>V4;样地3为V3>V4>V2>V1;样地4为V1>V2>V3>V4。当考虑未来的外部干扰时,各局域元江素馨种群年龄结构动态指数Vpi′分别为0.009 4、0.022 7、0.170 2及0.007 5,其增长性大小排序为样地3>样地2>样地1>样地4。各局域种群的年龄结构动态指数Vpi′较小,即种群为增长型,但趋向于稳定型。
2.3 种群数量动态预测
采用时间序列一次平均移动法进行了元江素馨种群动态预测,因所调查各局域元江素馨种群个体数量少,故不进行各局域种群的动态预测。以元江素馨各齡级现存活个体数为基础数据,采用一次移动平均法预测在未来2、4个龄级时间后各龄级个体数量,绘制元江素馨种群数量动态的时间序列预测图。由图4可知,元江素馨种群数量在未来2、4个龄级后呈增长趋势,如Ⅳ龄级原始个体数为36株,在2、4个龄级后分别增至49株、156株。Ⅴ龄级原始个体数为30株,在2、4个龄级后增至33株、61株。这表明元江素馨幼龄阶段个体数量充足,可为种群生长后期个体数提供补充。
3讨论与结论
3.1 元江素馨生境和群落学特征
自然界中,原生境所含自然资源和承载能力都是有限的,生境破碎化使种群原生境面积和容纳量减少,承载能力降低,种群的生存发展受限,甚至可能导致种群的消失。调查到元江素馨分布于海拔500~850 m范围内的干热河谷区,该区高温干燥、人为利用高、干扰强度大、生境常成片段化。且群落中乔木层物种少,林分郁闭度小,林内光照较强,以接受直射光为主,每日直射光照时间为8~10 h。而元江素馨种子萌发及幼苗生长需要一定的水分和荫蔽环境,显然如此恶劣的生存条件不利于其生存繁殖。
元江素馨各局域种群差异是由于环境差异及其自身生态适应能力造成的。元江素馨在发生发展过程中受多方面的环境因素影响,如人为干扰(砍伐、修路、放牧)、生物因子(物种丰富度、盖度)、地形地貌(坡度、坡向、坡位、岩石裸露程度)等。研究发现:(1)人为干扰不利于元江素馨种群的发展。海拔1 000 m以下的元江干热河谷是主要的农业耕作区,该区大量放牧、修建公路、近生活区存在盗木砍伐现象,一方面使生境片段化更加严重,减少元江素馨生存的原生生境,限制种群的发生发展空间;另一方面,牲畜践踏、啃食元江素馨幼苗、嫩枝嫩叶,使其生长发育受限,影响开花结实,人为砍伐使元江素馨幼苗丧失荫蔽环境而大量死亡或生长不良。(2)研究区物种丰富度低、盖度小,不利于元江素馨生长。植物有保水固土的作用,物种丰富度低、盖度小,一方面很难为元江素馨幼苗生长提供荫蔽环境,另一方面很难保证植物生长所需的水分、肥力等因子。(3)地形地貌对元江素馨种群发展产生一定影响。元江素馨多分布与西坡、西南坡的中部及中下部位,我们猜测是该区域地表水向下流动,冲刷地面,带走地表养分,林分中枯落物少、腐殖质少、土壤贫瘠、物种丰富度低、种间竞争相对较弱,可为元江素馨种群发展提供充足的空间,免去严酷的种内种间竞争等。同时,元江素馨努力争取适宜自身生存发展的空间,调查中也发现元江素馨可在石坑、石缝等生境中发展种群,可谓“夹缝中求生存”,展现出强大的生命力。而样地3元江素馨仅生存在牲畜无法啃食的灌木丛中,正是此种现象的真实写照。面对如此恶劣的生境,元江素馨形成了独特的生理适应机制以及具备了十分强大的生命力,如元江素馨叶片小而厚、根系异常发达,其地下部分长度常大于或等于地上部分高度、其种子仅在雨季萌发以及元江素馨可凭借顽强生命力在夹缝中求生等,正是元江素馨对干热河谷区长期适应的结果。
元江素馨主要生长于干热灌丛和干热性稀树灌木草丛中,群落中乔木层物种少而稀疏,灌木层、草本层物种较乔木层多而密。乔木层主要有三叶漆、清香木、厚皮树、余甘子、山合欢、心叶木,灌木层物种以疏序黄荆、小叶臭黄皮、元江素馨、老人皮、假虎刺、虾子花等为主,草本层物种以扭黄茅、旱茅、野骨草、黄背草、鬼针草等为主,藤本植物有毛叶鸡屎藤、镰叶西番莲、古钩藤等,数量较少,无附生植物,群落中物种组成以热带成分为绝对优势。元江素馨从幼龄阶段到中龄阶段死亡率高,我们推测其幼龄阶段因群落中草本在6-8月快速生长,为元江素馨幼苗生长提供相对荫蔽的环境,减少阳光直射,而使幼苗数量得以保存。但因草本层多为一年生植物,提供荫蔽环境的时间有限,乔木层物种稀疏、盖度小,无法为元江素馨继续生长提供相对有利的环境,又加上该区常年放牧,元江素馨被牲畜啃食而无法成苗。如此多的原因,致使元江素馨实生苗生长严重受限,幼苗期死亡率高。
3.2 元江素馨的种群结构特征
元江素馨各局域种群年龄结构呈两类变化趋势,增长型种群的Vpi′值均小于稳定型种群,故稳定型种群小生境更适合元江素馨生长。样地1、样地2、样地4种群呈现增长趋势,这与研究中种群存活曲线、种群结构类型及种群数量动态研究结果一致。其存活曲线表现为Deevey-Ⅲ型,与很多珍惜濒危植物种群如梵净山冷杉(李晓笑等,2011)、贵州槭(刘海燕等,2016)、巴山榧树(江波等,2018)的研究结果一致。种群Ⅰ龄级到Ⅱ龄级个体数减少较多,死亡率较高,一方面表明从幼龄阶段到中龄阶段,其种群生长发育受到限制,考虑是其自身生理机制造成,因此了解元江素馨生理工作机制是维持种群稳定发展、提高幼苗存活率的关键;另一方面考虑其高死亡率是极端生境造成的,幼龄阶段个体数较多,是其种子萌发率高还是产生种子数量多,其种子的萌发期是否与雨季有关等尚有待研究。当考虑未来外部干扰时样地2的Vpi′值远大于样地1、样地4,故样地2更适合元江素馨生长,出现此种现象的原因有待进一步研究。样地3种群呈现稳定趋势,存活曲线表现为Deevey-Ⅰ型,与新疆野苹果(田润炜等,2016)一致。种群Ⅰ龄级到Ⅱ龄级个体数减少较小,其幼苗能很好过渡到中龄阶段,同时调查中发现,此生境因相对荫湿且林分郁闭度较大,很多牲畜常在该生境下啃食及乘凉栖息,干扰严重,导致地表物种较少,元江素馨基本存留于牲畜无法啃食践踏的灌丛中,猜测这可能是其种群个体数量少特别是幼龄个体数量稀少的主要原因之一。
3.3 元江素馨种群动态趋势
元江素馨在干扰小的情况下,可实现种群的自我更新,种群数量缓慢增长。种群的壮大是一个漫长的过程,元江素馨增长速度较缓慢,若未来干扰强度变大且缺乏保护,种群将面临较高的死亡风险,甚至消失。因此,要保证元江素馨种群未来稳定发展并不断壮大,首先是了解其生物学特性及生殖繁育机制,促进母树结实,提高种子萌发率;其次是适当进行人工抚育,保证幼苗成活率,使幼苗继续稳定健康生长;最后是加强对其原生境的保护,减少人为破坏、牲畜践踏、生境流失等。
参考文献:
BAI PY, 1983. New taxa of Jasminum L. from Yunnan [J]. Acta Bot Yunnan, (1): 65-68. [白佩瑜, 1983. 云南素馨属新分类群 [J]. 云南植物研究, (1): 65-68.]
BAI PY, 1985. Notes on the genus Jasminum from Yunnan [J]. Acta Bot Yunnan, (4): 421-422. [白佩瑜, 1985. 云南素馨属植物的增补 [J]. 云南植物研究, (4): 421-422.]
CHENG SJ, TANG DQ, LIU QL, et al., 2012. Physiological characteristics of two species of Jasminum under salt stress [J]. J Fujian Coll For, (1): 33-38. [程淑娟, 唐东芹, 刘群录, 等, 2012. 盐胁迫下的2种素馨属植物生理特性 [J]. 福建林学院学报, (1): 33-38.]
CHEN XD, 1998. A study on the method of quantitative analysis for plant population and community structural dynamics [J]. Acta Ecol Sin, (2): 104-107. [陈晓德, 1998. 植物种群与群落结构动态量化分析方法研究 [J]. 生态学报, (2): 104-107.]
Chinese Botanical Society Editorial Board of Chinese Academy of Sciences, 1982. Flora Reipublicae Popularis Sinicae [M]. Beijing: Science Press, 61: 218. [中国科学院中国植物志编辑委员会,1982. 中国植物志 [M]. 北京:科学出版社, 61: 218.]
Flora of China Editorial Board, 1996. Flora of China [M]. Beijing: Science Press. Joint Publication by Missouri Botanical Garden Press, 15: 272-319. [中國科学院中国植物志编辑委员会,1996. 中国植物志 [M]. 北京:科学出版社. 密苏里植物园出版社联合出版, 15: 272-319.]
GE F, 2008. Modern Ecology [M]. Beijing: Science Press: 173. [戈峰, 2008. 现代生态学 [M]. 北京:科学出版社: 173.]
GUO QQ, LU J, LUO DQ, et al., 2010. Ecological characteristics of Sabina saltuaria population at the timberline ectone on the sunny slope of the Sejila Mountain [J]. J NW For Univ, (2): 15-18, 44. [郭其强, 卢杰, 罗大庆, 等, 2010. 西藏色季拉山阳坡林线方枝柏种群结构特征研究 [J]. 西北林学院学报, (2): 15-18,44.]
HAN L, WANG JQ, WANG HZ, et al., 2014. The population structure and dynamics of Populus euphratica at the upper reaches of the Tarim River [J]. Acta Ecol Sin, (16): 4640-4651. [韩路, 王家强, 王海珍, 等, 2014. 塔里木河上游胡杨种群结构与动态 [J]. 生态学报, (16): 4640-4651.]
JIANG B, ZHOU XR, SHANG J, et al., 2018. Population structure and dynamics of Torreya fargesii Franch, a plant endemic to China [J]. Acta Ecol Sin, (3): 1016-1027. [江波, 周先容, 尚进, 等, 2018. 中国特有植物巴山榧树的种群结构与动态 [J]. 生态学报, (3): 1016-1027.]
LI B, 2000. Ecology [M]. Beijing: Higher Education Press: 46. [李博, 2000. 生态学 [M]. 北京: 高等教育出版社: 46.]
LI XX,WANG QC, CUI GF, et al., 2011. Structure and dynamic characteristics of a wild population of endangered Abies fanjingshanensis [J]. Acta Bot Boreal-Occident Sin, (7): 1479-1486. ; [李晓笑, 王清春, 崔国发, 等, 2011. 濒危植物梵净山冷杉野生种群结构及动态特征 [J]. 西北植物学报, (7): 1479-1486.]
LIU GJ, ZHANG XM, ZHU JT, et al., 2011. Structure and dynamics of Haloxylon ammodendron population along the southeastern margin of the Zhunger Basin [J]. Acta Bot Boreal-Occident Sin, (6): 1250-1256. [刘国军, 张希明, 朱军涛, 等,2011. 准噶尔盆地东南缘梭梭种群结构与动态研究 [J]. 西北植物学报, (6): 1250-1256.]
LIU HY, YANG NK, ZOL TC, et al., 2016. Population structure and dynamics of the endemic plant Acer guizhouense in Guizhou Province [J]. Guihaia, 36(5):548-556. [刘海燕, 杨乃坤, 邹天才, 等, 2016. 贵州特有植物贵州槭的种群结构及动态研究 [J]. 广西植物, 36(5): 548-556.]
LIU JL, 2005. A new variety of Jasminum (Oleaceae) from Sichuan Province [J]. Bull Bot Res, (1):7-7. [刘建林, 2005. 四川素馨属(木犀科)一新变种 [J]. 植物研究, (1): 7-7.]
MAO BM, 1984. A revision of Jasminum in China [J]. Bull Bot Res, (1): 88-111. [缪柏茂, 1984. 中国素馨属的修订 [J]. 植物研究, (1): 88-111.]
PEI HG, WU JZ, CHEN SB, et al., 2013.Jasminum macrocarpum Merr. (Oleaceae), a newly recorded species from Vietnam [J]. J Trop Subtrop Bot, (4): 343-346. [裴洪光, 武进正, 陈世柏, 等, 2013越南木樨科植物新记录 [J]. 热带亚热带植物学报, (4): 343-346.]
SHEN SK, MA HY, WANG YH, et al., 2008. The structure and dynamics of natural population of the endangered plant Euryodendron excelsum H. T. Chang [J]. Acta Ecol Sin, (5): 2404-2412. [申仕康, 馬海英, 王跃华, 等, 2008. 濒危植物猪血木(Euryodendron excelsum H. T. Chang)自然种群结构及动态 [J]. 生态学报, (5): 2404-2412.]
TIAN RW, CAI XB, LIU LY, et al., 2016. Characteristics of the age structure and dynamics of Malus sieversii population [J]. Acta Bot Boreal-Occident Sin, (4): 811-817. [田润炜, 蔡新斌, 刘丽燕, 等, 2016. 新疆野苹果种群年齡结构特征与动态分析 [J]. 西北植物学报, (4): 811-817.]
WANG Z, HUANG RF, WANG LH, et al., 2009. Analysis on life table of natural Sabina vulgaris population in Muus Sandy Land [J]. J Desert Res, (1): 118-124. [王卓, 黄荣凤, 王林和, 等, 2009. 毛乌素沙地天然臭柏种群生命表分析 [J]. 中国沙漠, (1): 118-124.]
WEI ZX, ZHOU ZK, PAI PY, 1988. The pollen morphology of Jasminum L. [J]. Acta Bot Yunnan, (3): 271-279, 373-377. [韦仲新, 周浙昆, 白佩瑜, 1988. 素馨属植物花粉形态的研究 [J]. 云南植物研究, (3): 271-279, 373-377.]
XUE JH, 2006.Forest ecology [M]. Beijing: Forestry Press: 71. [薛建辉, 2006. 森林生态学 [M]. 北京:中国林业出版社: 71.]
YANG X, PENG H, LU SG, 2006. A review of phylogeny of Carlemanniaceae [J]. Bull Bot Res, (4): 397-401. [杨雪, 彭华, 陆树刚, 2006. 四角果科(Carlemanniaceae)的系统位置评述 [J]. 植物研究, (4): 397-401.]
YANG HX, ZHANG JT, WU B, et al., 2006. Point pattern analysis of Artemisia ordosica population in the Muus sandy land [J]. Acta Phytoecol Sin, (4): 563-570. [杨洪晓, 张金屯, 吴波, 等, 2006. 毛乌素沙地油蒿种群点格局分析 [J]. 植物生态学报, (4): 563-570.]
ZHANG WH, XU XB, ZHOU JY, et al., 2005. Population dynamics of endangered plant species Abies chensiensis [J]. Chin J Appl Ecol, (10): 1799-1804. [张文辉, 许晓波, 周建云, 等, 2005. 濒危植物秦岭冷杉种群数量动态 [J]. 应用生态学报, (10): 1799-1804.]
ZHANG XP, QIN H, YANG F, et al., 2014. Chemical consti-tuents of Jasminum giraldii and their antioxidant activity [J]. China J Chin Mat Med, (11): 2029-2033. [张修朋, 秦辉, 杨芳, 等, 2014. 黄素馨化学成分及其抗氧化活性 [J]. 中国中药杂志, (11): 2029-2033.]
ZHANG ZF, BIAN BL, TIAN XF, 2004. Advances in studies on chemical constituents and pharmacological activities of Jasminum [J]. Chin J Chin Mat Med, (12): 1209-1212. [张正付, 边宝林, 田秀峰, 2004. 素馨属植物化学成分及药理活性研究进展 [J]. 中国中药杂志, (12): 1209-1212.]
ZENG JQ, 2016. Population ecology of Alchornea davidii in karst areas of Chongqing [D]. Chongqing: Southwest University: 12-14. [曾嘉庆, 2016. 重庆喀斯特生境山麻杆种群生态学研究 [D]. 重庆: 西南大学: 12-14.]
