羊柴叶形态结构的地域分异特征及其与生态因子间的关系
2019-09-10杨洋赵杏花左合君
杨洋 赵杏花 左合君



摘要: 为探讨沙旱生植物的形态及组织结构对地域分异的响应,揭示植物形态、结构的地域分异规律。该研究以呼伦贝尔沙地、科尔沁沙地、浑善达克沙地及毛乌素沙地的羊柴(Hedysarum laeve)为对象,采用野外测量、石蜡切片技术及叶片解离技术,观察、分析了羊柴叶片形态及组织结构的地域分异特征,并就其形态、结构与生态因子间进行了相关性研究。结果表明:(1)随着干旱程度的加剧,羊柴叶片在形态上表现为变小、变厚,侧生小叶变长、变宽;在结构上表现为单位面积气孔数量减少,上、下表皮细胞变厚,栅栏组织越发达,栅栏细胞排列紧密,主脉维管束直径变大,疏导能力加强。(2)羊柴叶肉中存在大量的异细胞,具有抵御干旱及贮藏等功能,是对干旱环境长期适应的体现。(3)生长季降水量、年均降水量及年均蒸发量是形成羊柴小叶形态、结构地域分异的主要环境因子。以上结果表明植物对地域分异的响应不仅体现在外部形态上,在组织结构上同样产生相应的分异特征,而水热条件则是产生分异的主要因素。
关键词: 羊柴叶, 形态特征, 结构特征, 异细胞, 地域差异
中图分类号: Q944.56文献标识码: A文章编号: 1000-3142(2019)09-1233-10
Abstract: In order to explore the response of morphology and structure of xerophytes to regional differentiation, leaf morphology and structure of Hedysarum laeve in Hulunbuir, Horqin, Hunshandake and Mu Us sandy lands were observed and analyzed by field measurement, paraffin section technology and leaf dissociation technology. The relationship among morphology, structure and ecological factors was also studied. The results were as follows: (1) With drought intensifies, the morphological characteristics of H. laeve leaf were smaller, thicker, lateral lobules longer and wider; stomatal number per unit area decreased, upper and lower epidermal cells became thicker, palisade tissue became more developed, palisade cells arranged closely, diameter of main vascular bundle became larger, and dredging ability was strengthened. (2) H. laeve mesophyll contained a large number of idioblast, which had the functions of drought-resisting and storage, and reflected the long-term adaptation of drought environment. (3) The precipitation in growing season, annual average precipitation and annual average evaporation were the main environmental factors for the formation of H. laeve lobule morphology and regional structure differentiation. This indicates that the response of plants to regional differentiation is not only reflected in the external morphology, but also in the organizational structure, and the hydrothermal conditions are the main factors.
Key words: leaf of Hedysarum laeve, morphological characteristics, structure characteristics, idioblast, regional difference
地域分异是地理环境各组成要素及其构成的自然综合体在地表沿一定方向规律性分异的现象,支配这种分异现象的客观规律为地域分异规律(李少娟等,2007)。不同地域分布的植物,由于地带性而产生的水热条件差异,在形态、结构上表现出明显差异(李正理,1981;贺金生等,1994)。被子植物的叶是进行光合作用的主要器官,为植物的生长提供所需的营养物质,对植物的生长发育起决定性作用;同时,叶片是植物对环境变化较为敏感的器官,其形态、结构特征更能体现其对地域分异的适应(李琪,2017;刘静等,2011)。
生长在同一地域的不同种植物,其形态、结构特征会表现出趋同现象;与此相反,生长在地域差异非常大的同种植物,它們的形态及结构特征一般表现出较明显的趋异现象(陈庆诚等,1961)。羊柴(Hedysarum laeve),又名塔落岩黄芪,豆科岩黄芪属旱生灌木,是我国沙区广泛分布的固沙树种(马毓泉,1994)。为了探究植物形态结构对地域分异的响应,自东向西选取具有明显地域差异的呼伦贝尔沙地、科尔沁沙地、浑善达克沙地及毛乌素沙地作为采样区,以四个样区内共有植物羊柴叶片作为研究对象,观测其在不同地区的形态、结构特征。目前,对羊柴的研究主要集中在叶表皮微形态、解剖结构上(金洪和周丽霞,1990;胡月楠等,2008;李琪,2017),但对其叶片形态结构在不同地域的差异性少有报道。本研究旨在探讨羊柴叶形态及组织结构的地域分异,了解羊柴的分布及其生态适应过程,为进一步揭示旱生植物形态、结构和功能对地域差异的适应机制提供依据。
1采样地概况
呼伦贝尔沙地位于大兴安岭针叶林区,属温带大陆性气候,≥10 ℃积温为1 800~2 200 ℃;科尔沁沙地位于西辽河流域干草原区,属温带大陆性季风气候,四季分明,≥10 ℃以上积温为3 160 ℃;浑善达克沙地位于内蒙古高原干草原与荒漠草原的过渡区,属中温带干旱大陆性气候,≥10 ℃以上积温为1 970 ℃;毛乌素沙地位于鄂尔多斯高原干草原与荒漠草原过渡区,属温带大陆性季风气候,降水年际变率大,多雨年为少雨年2~4倍,常发生旱灾和涝灾,且旱多于涝,≥10 ℃以上积温为2 945 ℃。具体采样地环境参数如表1所示。
2材料与方法
2.1 野外数据测量
研究材料于2017年8月测量及采集。在各样地分别选取5丛冠幅大小相近的灌丛,每个灌丛随机各选取40片新鲜的羊柴顶端小叶及侧生小叶,用电子数显卡尺测量叶片长、宽及叶厚,并计算叶片的长宽比。
2.2 试验材料采集
在各样地分别选取5丛冠幅大小相近的灌丛,每个灌丛采集无病害的当年生枝条羽状复叶顶端小叶(简称顶端小叶,下同)和侧生小叶各40片,用清水冲洗叶片表面的泥土后放入FAA固定液中保存。
2.3 石蜡制片及指标测定
叶片结构研究采用常规石蜡切片法制片(横切),切片厚度为14 μm,番红-固绿对染,中性树胶封片,Motic显微镜进行观察,Motic DigilabⅡ拍照系统进行拍照,Motic Images Plus 2软件进行指标的测量。主要测量指标有上、下表皮细胞厚,靠近上表皮一侧栅栏组织厚(简称上栅栏组织,下同), 靠近下表皮一侧栅栏组织厚 (简称下栅栏组织,下同),主脉直径。
2.4 气孔观测
叶片气孔采用解离法,解离法是一种非切片制片法,在固定液中取出羊柴小叶,用蒸馏水冲洗干净,放入冰醋酸∶双氧水(30%)=1∶1的解离液中,直至叶片变成半透明状,用毛笔轻轻刷开叶片的上或下表皮,酒精梯度脱水,固绿染色,中性树胶封片。主要测量指标有小叶的上、下表皮气孔长、宽、长宽比及单位面积气孔数量。气孔密度是指分布范围内的单位面积气孔个数。气孔密度=固定面积内气孔器个数/固定面积(李琪,2017)。
2.5 数据处理
数据统计分析采用单因素方差分析(One-way ANOVA)和相关分析。
3结果与分析
3.1 叶形态特征的地域分异
羊柴顶端小叶长度为15.36~21.57 mm(表2),采自毛乌素沙地与浑善达克沙地、科尔沁沙地和呼伦贝尔沙地的羊柴小叶无显著差异,浑善达克沙地与科尔沁沙地和呼伦贝尔沙地有显著差异,科尔沁沙地与呼伦贝尔沙地无显著差异。从顶端小叶宽度和长宽比来看,宽度为3.51~4.47 mm,比值为4.23~4.93,采自四个沙地的羊柴小叶无显著差异。侧生小叶长度为11.18~14.85 mm,采自毛乌素沙地与浑善达克沙地和科尔沁沙地的羊柴小叶无显著差异,与呼伦贝尔沙地有显著差异,浑善达克沙地与科尔沁沙地无显著差异,与呼伦贝尔沙地有显著差异,科尔沁沙地与其余三个沙地无显著差异。侧生小叶宽度为2.93~4.27 mm,采自毛乌素沙地与浑善达克沙地和科尔沁沙地的羊柴小叶无显著差异,与呼伦贝尔沙地有显著差异,浑善达克沙地与其余三个沙地无显著差异,科尔沁沙地与其余三个沙地无显著差异。侧生小叶长宽比为3.32~3.91,采自四个沙地的羊柴小叶无显著差异。小叶厚度1.77~2.51 mm,采自四个沙地的羊柴小叶均有显著差异。 叶片变小、变厚,既可减少叶片的蒸腾面积,又能有效提高其贮水性,是植物从形态上对地域差异的适应性做出的改变(Wells et al.,1991)。本研究中,羊柴小叶长宽比没有明显的变化, 由此可以说明叶形保持稳定。而从小叶厚度来看,具有明显的地域分异,其中科尔沁沙地的水分条件最好,小叶最薄;浑善达克沙地的水分条件最差,小叶最厚。
3.2 叶结构特征的地域分异
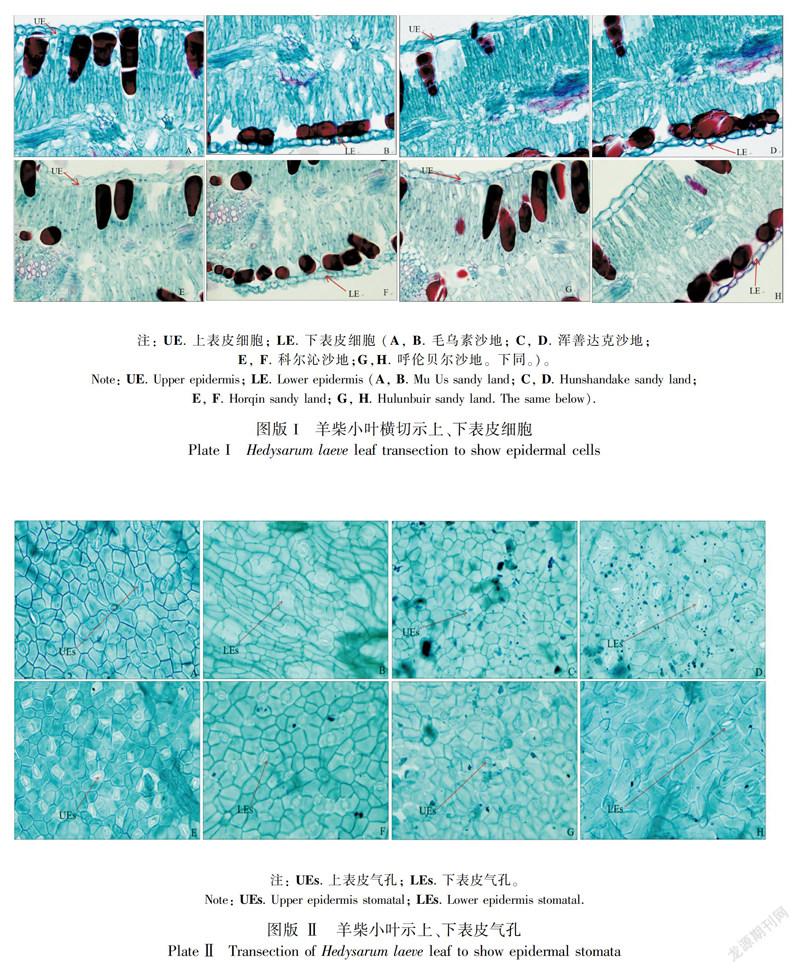
双子叶植物叶由表皮、叶肉和叶脉构成。羊柴小叶表皮由一层细胞构成,细胞排列紧密,无胞间隙;上下表皮均有气孔分布且有气孔腔(图版I)。采自于不同地域的羊柴小叶上表皮厚度为77.56~101.45 μm(表3),毛乌素沙地与浑善达克沙地和呼伦贝尔沙地有显著差异,与科尔沁沙地无显著差异,浑善达克沙地与呼伦贝尔沙地、科尔沁沙地有显著差异,呼伦贝尔沙地与其余三个沙地均有显著差异。下表皮细胞厚度为71.76~88.41 μm,采自毛乌素沙地与浑善达克沙地和呼伦贝尔沙地的羊柴小叶有显著差异,与科尔沁沙地无显著差异,浑善达克沙地与呼伦贝尔沙地无显著差异,与科尔沁沙地有显著差异,科尔沁沙地与呼伦贝尔沙地有显著差异。密集的表皮细胞和平直的细胞壁,可以有效地控制水分的散失(白潇等,2013)。本研究中,内蒙古四个沙地的干旱程度由强到弱的顺序依次是浑善达克沙地、呼伦贝尔沙地、毛乌素沙地、科尔沁沙地,分布在四个沙地上的羊柴小叶上、下表皮厚度均是科尔沁沙地的最薄,浑善达克沙地的最厚,这是植物的组织结构对地域差异适应的具体体现。
羊柴小叶上表皮单位面积气孔数量为25.71~40.72 个·mm-2(表3),采自毛乌素沙地的羊柴小叶与科尔沁沙地和呼伦贝尔沙地无显著差异,与浑善达克沙地有显著差异,浑善达克沙地与科尔沁沙地和呼伦贝尔沙地有显著差异,科尔沁沙地与呼伦贝尔沙地无显著差异。从小叶上表皮气孔长度、宽度和长宽比来看,长度为75.05~77.10 μm,宽度为47.05~54.10 μm,比值为1.41~1.63,采自四个沙地的羊柴小叶均无显著差异。羊柴小叶下表皮单位面积气孔数量为42.93~61.40 个·mm-2,采自毛乌素沙地的羊柴小叶与浑善达克沙地、科爾沁沙地和呼伦贝尔沙地有显著差异,浑善达克沙地与科尔沁沙地和呼伦贝尔沙地有显著差异,科尔沁沙地与呼伦贝尔沙地无显著差异。小叶下表皮气孔长度为75.25~79.43 μm, 采自毛乌素沙地的羊柴小叶与科尔沁沙地和呼伦贝尔沙地无显著差异,与浑善达克沙地有显著差异,浑善达克沙地与科尔沁沙地和呼伦贝尔沙地有显著差异,科尔沁沙地与呼伦贝尔沙地无显著差异。从羊柴小叶下表皮气孔宽度和长宽比来看,宽度为48.90~59.00 μm,比值为1.28~1.61,采自四个沙地的羊柴小叶均无显著差异。气孔器越小,植物对水分利用率越大,且其多分布于下表皮可以有效地使植物与外界进行气体交换,减少蒸腾作用(赵庆芳等,2006)。本研究中,随着降水量的减少,羊柴小叶气孔数量也随之减少,这是植物为降低蒸腾而形成的结构差异。
双子叶植物的叶肉一般由栅栏组织和海绵组织构成,对于旱生植物而言,栅栏组织发达,海绵组织退化。羊柴小叶栅栏组织非常发达,海绵组织退化甚至消失,使栅栏组织呈双栅型,上栅栏组织细胞呈长柱型,排列紧密,由3~5层细胞组成;下栅栏组织细胞呈不规则长柱型,排列疏松,由2~3层细胞组成(图版Ⅲ)。羊柴小叶上栅栏组织厚度为628.96~917.42 μm,采自不同地域的叶均有显著差异。下栅栏组织厚度为421.80~634.90 μm,采自毛乌素沙地与科尔沁沙地的羊柴小叶无显著差异,与浑善达克沙地和呼伦贝尔沙地有显著差异;采自浑善达克沙地和呼伦贝尔沙地的羊柴小叶与其余地域的叶有显著差异;采自科尔沁沙地的羊柴小叶与呼伦贝尔沙地有显著差异。发达的栅栏组织可减小甚至避免光照对叶肉细胞的损害,有利于植物在干旱条件下生长(郭婧宇等,2017)。本研究中,随着干旱程度的加剧,羊柴小叶的栅栏组织变厚、层数增加且细胞变长。
本研究结果发现,在不同地域采集的羊柴叶横切中,靠近上表皮一侧均有被番红染色的长柱状薄壁细胞分布(长径垂直于上表皮细胞);而在靠近下表皮一侧也存在着一层被番红染色的圆柱状的薄壁细胞(长径平行于下表皮细胞),这些薄壁细胞着色较深,含有丰富的后含物,具有贮藏功能,是植物对环境适应过程中形成的异细胞(图版Ⅲ)。
羊柴小叶主脉的维管束鞘由较大的薄壁细胞组成,形状规则且排列整齐;主脉维管束呈扇形,木质部有口径不同的数列导管,且口径较大;韧皮部韧皮纤维比较发达(图版IV)。羊柴小叶主脉直径为719.33~789.37 μm(表4),采自毛乌素沙地与科尔沁沙地的羊柴小叶无显著差异,毛乌素沙地、浑善达克沙地、呼伦贝尔沙地彼此之间有显著差异;科尔沁沙地、浑善达克沙地、呼伦贝尔沙地彼此之间有显著差异。叶片中维管组织越发达,则运输能力越强,植物的光合效率越高,这是植物对干旱条件的适应(白潇等,2013;杨九艳等,2005)。本研究结果表明,随着降水量减少,羊柴小叶主脉维管束直径越发达,运输能力越强。
3.3 叶形态、结构与环境因子的相关性
对羊柴小叶形态、结构与环境因子进行相关性分析(表5)结果表明,羊柴侧生小叶长与年均蒸发量呈显著负相关关系;顶端及侧生小叶长宽比与年均降水量呈显著正相关关系;叶厚,上、下表皮厚和上、 下栅栏组织厚与年均降水量呈显著负相关关系;上表皮气孔长与生长季降水量呈极显著负相关关系;侧生小叶宽与生长季降水量呈显著负相关关系。这说明羊柴小叶的形态、结构的改变受降水量、温度及蒸发量等气象因子的影响,而生长季降水量、年均降水量和年均蒸发量是小叶形态结构发生变化的主要影响因子,随着降水量的减少,小叶变的小而厚,表皮细胞及栅栏组织变厚,主脉直径变大,单位面积气孔数量减少,采自浑善达克沙地的小叶最厚,表皮细胞及栅栏组织最厚,主脉直径最大,单位面积气孔数量最少。
4讨论
叶片是植物对地域性较为敏感的器官,其形态结构特征很容易受到地域性的影响,表现出叶片外部形态及内部结构差异性以及对地域差异的适应性(赵庆芳等,2006;张文辉等,2004;陈健辉等,2018)。本研究发现,浑善达克沙地水分条件最差,羊柴小叶最厚;科尔沁沙地水分条件最好,羊柴小叶最薄;呼伦贝尔沙地和毛乌素沙地水分条件居中,羊柴小叶厚度也位居于中,这一现象符合相关分析得出的结果,即羊柴叶厚与年均降水量呈显著负相关关系。水分是植物在干旱生境中的限制性因子,植物为了减少蒸腾并贮藏更多水分,在形态、结构上表现为叶片变得小而厚,在减少蒸腾表面积的同时增加贮水组织的比例;同时,以减少气孔数量来降低蒸腾(施海燕等,2009;马全林等,2008),本研究结果与之相一致。由此可见,羊柴适应不同地域的气候、环境差异,在形态、结构上表现明显的地域分异。
异细胞是植物组织中呈现出的特殊植物细胞,较为常见的是在逆境生长的植物叶片中具有抗逆功能的异细胞,主要分布在叶肉栅栏组织靠近上、下表皮侧等部位。酸模属7种植物叶片表皮中存在体积明显大于普通表皮细胞的异细胞,形状为球形或椭圆形(李波等,2008)。对16种高山植物叶片进行有解剖学研究发现,在叶肉的栅栏组织靠近上表皮的位置、海绵组织以及上、下表皮存在异细胞,这些异细胞可能与高山植物的抗逆性有关,具有抵御干旱、强太阳辐射等作用,但细胞成分不明(祁如虎等,1998)。本研究中,不同地域羊柴小叶横切中靠近上、下表皮均有被番红染色的薄壁细胞分布,这些胞壁细胞明显较周围其他细胞大,含有丰富細胞贮藏物质,四个沙地中,浑善达克沙地最旱,科尔沁沙地最湿润,其余二者居中,四个沙地羊柴叶肉中的异细胞数量与其干旱程度的变化是一致的,是植物对干旱、风沙等逆境适应的体现。
降水、温度及蒸发量等气象因子对沙棘等旱生植物叶片的气孔密度、栅栏组织和海绵组织的分化有一定程度的影响,随着干旱程度增加,植物叶面积减小,气孔密度减小,表皮细胞增厚,栅栏组织发达,海绵组织退化甚至消失,主脉发达等变化是植物在解剖结构上对水分不足给出的响应(王怡,2003;何涛等,2007;陈庆诚等,1961)。本研究中,科尔沁沙地的羊柴小叶解剖特征表现为单位面积气孔数量最多,上、下表皮细胞和上、下栅栏组织最薄,主脉直径最小;浑善达克沙地羊柴小叶单位面积气孔数量最少,上、下表皮细胞和上、下栅栏组织最厚,主脉直径最大。这些组织结构的变化,与地域环境的水热变化规律一致,说明随着地域环境的分异可以使植物的形态发生分异,组织结构也会产生相应的地域分异。
5结论
(1)呼伦贝尔沙地、科尔沁沙地、浑善达克沙地及毛乌素沙地在自然分布上具有明显的地域性,其共有植物羊柴小叶在形态上形成明显的地域差异,随着干旱程度的加剧,羊柴小叶变小、变厚,侧生小叶变长、变宽,但长宽比没有明显的变化,叶形保持稳定。(2)从羊柴小叶的组织结构水平分析,随着干旱程度的加剧,表皮细胞变厚,栅栏组织变厚、层数增加且栅栏组织细胞变长,主脉直径变大,单位面积气孔数量减少。分布在不同地域的羊柴叶肉中均存在异细胞,其中上表皮一侧异细胞均明显大于下表皮一侧,这些薄壁细胞着色比较深,含有丰富的细胞后含物。(3)生长季降水量、年均降水量和年均蒸发量对羊柴小叶形态建成及组织结构的形成具有显著影响,分布在四个沙地的羊柴小叶在形态及结构上形成的分异特征主要源于水分条件的差异。
参考文献:
BAI X, LI Y, SU SP, et al., 2013. Response of leaf anatomical characteristic of Nitraria tangutorum Bobr. from different populations to habitats [J]. Acta Bot Boreal-Occident Sin, 33(10): 1986-1993. [白瀟, 李毅, 苏世平, 等, 2013. 不同居群唐古特白刺叶片解剖特征对生境的响应研究 [J]. 西北植物学报, 33(10):1986-1993.]
CHEN QC, SUN YW, ZHANG GL, 1961. A preliminary study on the morphology and anatomy of the dominants of the vegetations distributing along the middle and low drainage basin of the Shor-ler River based on the view point of ecology [J]. J Lanzhou Univ, (3): 61-96. [陈庆诚, 孙仰文, 张国樑, 1961. 疏勒河中、下游植物群落优势种生态-形态, 解剖特性的初步研究 [J]. 兰州大学学报, (3):61-96.]
CHEN JH, MIAO SY, HUANG HF, et al., 2018. Comparative study on leaf structures of five mangrove plants [J]. Guihaia, 38(5): 655-664. [陈健辉, 缪绅裕, 黄惠芳, 等, 2018. 五种红树植物叶片结构的比较研究 [J]. 广西植物, 38(5):655-664.]
GUO JY, ZHAO XH, ZUO HJ, 2017. Environmental different characteristic study on anatomical structure of Caragana korshinskii leaflet [J]. Acta Bot Boreal-Occident Sin, 37(12): 2396-2400. [郭婧宇, 赵杏花, 左合君, 2017. 柠条锦鸡儿小叶解剖结构的环境分异性研究 [J]. 西北植物学报, 37(12):2396-2400.]
HU YN, HE KN, GONG YX, et al., 2008. Leaf water potential of Hedysarum mongolicum in Kubuqi Desert of Inner Mongolia [J]. J W Chin For Sci, 3(37):66-71. [胡月楠, 贺康宁, 巩玉霞, 等, 2008. 库布齐沙地杨柴叶水势的研究 [J]. 西部林业科学, 3(37):66-71.]
HE T, WU XM, JIA JF, et al., 2007. Research advances in morphology and anatomy of alpine plants growing in the Qinghai-Tibet plateau and their adaptations to environments [J]. Acta Ecol Sin, 7(6): 2574-2583. [何涛, 吴学明, 贾敬芬, 等, 2007. 青藏高原高山植物的形态和解剖结构及其对环境的适应性研究进展 [J]. 生态学报, 27(6):2574-2583.]
HE JS, CHEN WL, WANG XL, et al., 1994. Morphological and anatomical factures of Quercus section Suber and its adaptation to the ecological environment [J]. Acta Phytoecol Sin, 18(3): 219-227. [贺金生, 陈伟烈, 王勋陵, 等, 1994. 高山栎叶的形态结构及其与生态环境的关系 [J]. 植物生态学报, 18(3):219-227.]
JIN H, ZHOU LX, 1990. Study on anatomical structure of vegetative organs of Hedysarum laeve and Hedysarum scoparium [J]. Inner Mongolia Pratac, (1): 53-57. [金洪, 周丽霞, 1990. 羊柴和花棒营养器官解剖结构的研究 [J]. 内蒙古草业, (1): 53-57.]
LI Q, 2017. Comparative study on leaf structure characteristics of 15 species of Leguminosae [D]. Hohhot: Inner Mongolia Agricultural University. [李琪, 2017. 豆科15种植物叶片结构特征比较研究 [D]. 呼和浩特: 内蒙古农业大学.]
LIU J, ZHAO HY, MA M, 2011. Anatomy of stem and leaf in different populations of Capparis spinosa in China [J]. Guihaia, 31(6): 735-740. [刘静, 赵红艳, 马淼, 2011. 旱生植物刺山柑不同种群茎叶的解剖学研究 [J]. 广西植物, 31(6):735-740.]
LI ZL, 1981. Morphology and structure of xerophytes [J]. Bull Biol, (4): 9-12. [李正理, 1981. 旱生植物的形态和结构 [J]. 生物学通报, (4):9-12.]
LI B, ZHANG WG, CHEN SF, et al., 2008. Comparative ana-tomy of the leaves of Rumex in Jiangxi [J]. J Wuhan Bot Res, (5): 443-449. [李波, 张文根, 陈少风, 等, 2008. 江西产酸模属植物叶片比较解剖学研究 [J]. 武汉植物学研究, (5):443-449.]
LI SJ, HE DM, ZHANG YP, 2007. Temporal and spatial variation of precipitation and its regional differentiation in longitudinal ridge-valley region [J]. Chin Sci Bull, 52(Suppl.): 51-63. [李少娟, 何大明, 张一平, 2007. 纵向岭谷区降水量时空变化及其地域分异规律 [J]. 科学通报, 52(增刊):51-63.]
MA YQ, 1994. Inner Mongolia Flora [M]. 2nd Ed. Huhhot: Inner Mongolia People Publishing House, 3: 339-340. [ 马毓泉, 1994. 内蒙古植物志 [M]. 2版. 内蒙古人民出版社, 3:339-340.]
MA QL, LIU SZ, YAN ZZ, et al., 2008. Drought resistant characteristics of wild Allium mongolicum [J]. Pratac Sci, 25(6): 56-61. [马全林, 刘世增, 严子柱, 等, 2008. 沙葱的抗旱性特征 [J]. 草业科学, 25(6):56-61.]
WELLS OO, SWITZER GL, SCHMIDTLING RC , 1991. Geographic variation in Mississippi Loblolly pine and sweetgum [J]. Silv Gen, 40:105-119.
QI RH, LI YZ, FAN J, 1998. The idioblasts and their ecological significance in leaves of 16 alpine plants [J]. Acta Bot Boreal-Occident Sin, 18(2):119-125. [祁如虎, 李有忠, 范建, 1998. 16种高山植物叶片内的异细胞及其生态学意义 [J]. 西北植物学报, 18(2):119-125.]
SHI HY, WANG YF, WANG JH, et al., 2009. Studies on the anatomical structures of leaves of seven Saussurea DC. species on the Qinghai-Tibetan plateau with different altitude [J]. Bull Bot Res, 29(3): 289-294. [施海燕, 王一峰, 王剑虹, 等, 2009. 青藏高原不同海拔地7种风毛菊属植物叶片解剖结构的研究 [J]. 植物研究, 29(3):289-294.]
WANG Y, 2003. Contrasts and observations of dissecting constructures of three drought-resistant plant leaf slices [J]. J Sichuan For Sci Technol, 24(1): 64-67. [王怡, 2003. 三种抗旱植物叶片解剖结构的对比观察 [J]. 四川林业科技, 24(1):64-67.]
YANG JY, YANG J, YANG MB, et al., 2005. Studies on the ecological adaptability of the genus Caragana on the ordos plateau [J]. J Arid Land Resour Environ, 19(3): 175-179. [杨九艳, 杨劼, 杨明博, 等, 2005. 鄂尔多斯高原锦鸡儿属植物叶的解剖结构及其生态适应性 [J]. 干旱区资源与环境, 19(3):175-179.]
ZHAO QF, MA RJ, DU GZ, et al., 2006. Ecological signification of the leaf structure of three species of Ligularia in different altitudes [J]. J Lanzhou Univ (Nat Sci Ed), 42(1): 33-39. [赵庆芳, 马瑞君, 杜国桢, 等, 2006. 不同海拔三种橐吾属植物叶结构的适应意义 [J]. 兰州大学学报(自然科学版), 42(1):33-39.]
ZHANG WH, LU T, MA KM, et al., 2004. Analysis on the environmental and spatial factors for plant community distribution in the arid valley in the upper reach of Minjiang River [J]. Acta Ecol Sin, 24(3): 552-559. [張文辉, 卢涛, 马克明, 等, 2004. 岷江上游干旱河谷植物群落分布的环境与空间因素分析 [J]. 生态学报, 24(3):552-559.]
