从江香猪源干扰素α2、β在生菜中的瞬时表达及其对PRRSV复制抑制作用的研究
2019-09-04温贵兰张升波杨佰启李昌红张喜懿龚新勇陈彦希
田 浪,温贵兰*,张升波,杨佰启,李昌红,徐 丽,陈 广,张喜懿,龚新勇,陈彦希,文 明
(1.贵州大学动物科学学院/预防兽医实验室,贵州贵阳550025;2.贵州省动物生物制品工程技术研究中心,贵州贵阳550016)
目前我国养猪业已经形成较大的规模,但由于高密度的养殖、环境的恶化和抗生素的滥用等因素,猪感染的病毒病也越来越多。猪繁殖与呼吸综合征(Porcine reproductive and respiratory syndrome,PRRS)是由PRRS病毒引起的猪一种繁殖障碍和呼吸系统高度接触性传染病,被认为是影响养猪业健康发展的重要疾病之一,目前尚无特效药治疗PRRS,因此研制高效、经济、安全的抗PRRSV新型药物制剂显得非常重要。
干扰素(Interferon,IFN)是在特定的诱生剂作用下,由细胞产生的一种具有高度生物学活性的糖蛋白,具有广谱抗病毒活性[1]。IFN本身并非直接抗病毒物质,不直接与靶分子作用,而首先要与靶细胞表面的特异性受体相结合,通过信号传递,引发一系列特定的生化反应,刺激细胞内多种效应蛋白质分子合成,从而发挥IFN的活性功能[2]。猪IFN-α、猪IFN-β属于IFN-I,具有重要的抗病毒生物学活性。已有研究人员在毕赤酵母或杆状病毒中表达了猪IFN-α,表达蛋白对PRRSV、猪传染性胃肠炎病毒、猪流行性腹泻病毒、猪圆环病毒2型具有抑制作用[3-5];在毕赤酵母表达了猪IFN-β,表达蛋白对伪狂犬病毒、PRRSV、水泡性口炎病毒具有抑制作用[6-8]。
从江香猪主产于贵州省从江县,属于中国稀有微型的地方优良品种猪,其具有基因纯合、近交不退化、适应性强和抗病力强等优良特点,是猪相关疾病研究的理想动物模型,有着较高的研究价值[9]。温贵兰等报道从江香猪源IFN-α2核苷酸序列与猪源IFN-α2核苷酸序列同源性为98.7%,同时存在7处碱基的变异,前者推导的氨基酸序列与后者存在4处差异;从江香猪IFN-β基因核苷酸序列与其它猪源IFN-β基因同源性为99.5%~100%,其氨基酸序列与巴马猪、梅山猪IFN-β的氨基酸序列同源性均为100%,但与贵州白香猪IFN-β的氨基酸序列同源性为99.5%,存在E43Q、K73R和C16R 3处氨基酸差异[10-11]。到目前为止,尚未见融合从江香猪源IFN-α2、IFN-β于生菜中瞬时表达并对瞬时表达蛋白进行抗病毒活性研究的相关报道。本实验构建了含从江香猪源 IFN-α2、IFN-α2β、IFN-β 的植物表达载体并转化根癌农杆菌LBA4404,采用根癌农杆菌渗透法感染生菜,对瞬时表达的从江香猪源IFN-α2、IFN-α2β、IFN-β 蛋白进行抗 PRRSV 活性研究,以期为从江香猪源IFN抗PRRSV新复合型药物的开发利用提供参考依据。
1 材料与方法
1.1 主要实验材料 含从江香猪源IFN-α2、IFN-β、IFN-α2β的质粒pUCm-CJpoIFN-α2、pUCm-CJpoIFN-β、pUCm-CJpoIFN-α2β 以 及 PRRSV GZBJ201712 株 、Marc 145细胞、PK15细胞均由本实验室保存;新鲜生菜购自贵州省花溪区某农贸市场。
根癌农杆菌LBA4404感受态细胞购自上海维地生物技术有限公司;无内毒素质粒小提中量试剂盒、TRIzol试剂、SmaⅠ、XbaⅠ、DL2000 DNA Marker、SYBR Premix ExTaqⅡ、HiFiScript cDNA Synthesis Kit试剂盒、β-葡萄糖苷酶(GUS)染液试剂盒均购自宝生物工程(大连)有限公司;λ DNA/HindⅢ购自天根生化科技有限公司;乙酰丁香酮、2-(N-吗啡啉)乙磺胺(MES)均购自北京索莱宝科技有限公司;猪IFN-α ELISA试剂盒购自武汉华美生物工程有限公司;猪IFN-β ELISA试剂盒购自武汉伊莱瑞特生物科技股份有限公司;BCA蛋白浓度测定试剂盒、2×EsTaqDNA聚合酶购自康为世纪生物科技有限公司;植物双元表达载体pBI121购自上海继和生物科技有限公司。
1.2 引物的设计与合成 参考GenBank登录的猪IFN-α2 基因序列(NM_001130219)、IFN-β 基因序列(GQ415073)、PRRSV全基因组序列(EF112445)、β-actin基因序列(NM_001199954),利用软件设计扩增 IFN-α2、IFN-β、IFN-α2β、N 基因、β-actin 的引物序列(表1)。引物均由上海生工生物工程技术服务有限公司合成。下划线为酶切位点。
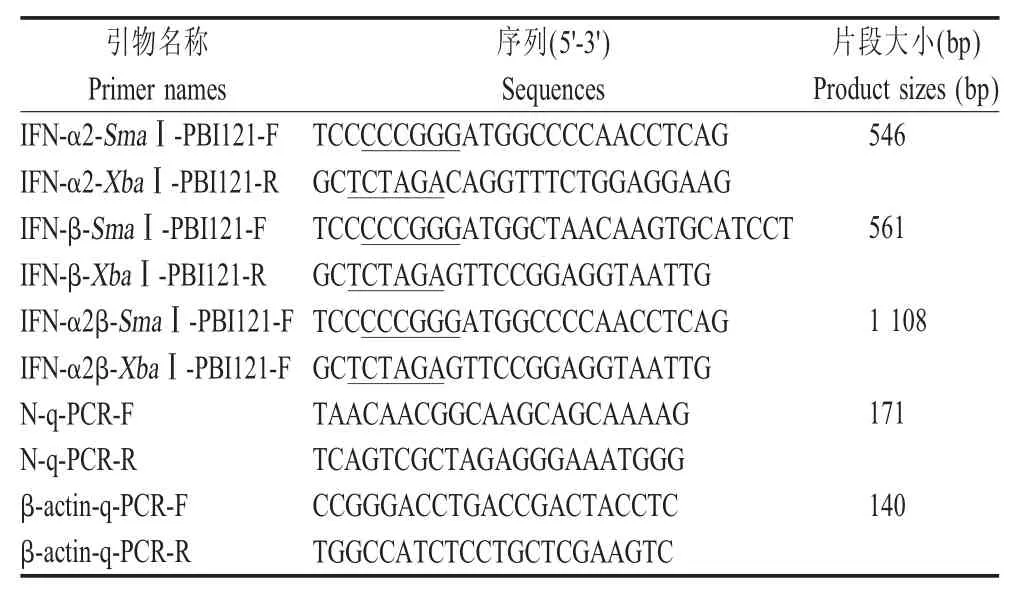
表1 本研究所用引物Table 1 The sequences of the primers used in this study
1.3 含从江香猪源各干扰素植物表达载体的构建与鉴定 采用设计扩增各基因的引物,分别以pUCm-CJpoIFN-α2、pUCm-CJpoIFN-β、pUCm-CJpoIFN-α2β质粒为模板对 IFN-α2、IFN-β、 IFN-α2β 基因片段进行PCR扩增,PCR扩增条件:94℃2 min;94℃30 s、55℃30 s、72℃1 min,共35个循环;72℃10 min。扩增产物经1.2%琼脂糖凝胶电泳检测后回收纯化。经SmaⅠ和XbaⅠ双酶切后连接至经相同双酶切的植物表达载体pBI121中,构建重组质粒pBI121-IFN-α2、pBI121-IFN-β、pBI121-IFN-α2β。将各质粒包括空质粒pBI121-Vec分别转入根癌农杆菌LBA4404中,菌液PCR筛选阳性重组菌。对阳性重组菌扩大培养,经无内毒素质粒小提中量试剂盒提取各质粒经PCR、双酶切鉴定后由英潍捷基(上海)贸易有限公司测序鉴定。
1.4 从江香猪源干扰素浸染液的制备 取含pBI121-IFN-α2、 pBI121-IFN-β、 pBI121-IFN-α2β、pBI121-Vec质粒的根癌农杆菌LBA4404菌液,28℃振荡培养至OD600nm值为0.6~0.8时转移至的10 mL离心管中,4 000 r/min室温离心5 min,分别收集各菌体沉淀,利用液体MS培养基重悬至原体积,加入2 mL 50 mmol/L乙酰丁香酮及10 mL 0.5 mol/L MES,使其终浓度分别为200 μmol/L和10 mmol/L,即为浸染液。
1.5 根癌农杆菌介导从江香猪源干扰素的真空渗透瞬时表达 新鲜生菜去除损伤叶片,用自来水漂洗2~3次,再用无菌水漂洗2~3次后超净工作台内风干,将用无菌针头刮伤的叶片放入1.4制备的浸染液中,在0.08 mbar气压下连续抽真空40 min,关掉真空泵,快速排气。取出浸染好的刮伤生菜叶片用无菌水漂洗2~3次,滤干后放入铺有无菌吸水纸的培养皿中,置于植物光照培养箱中,在22℃每天光照16 h,光照强度为2000 lx。培养48 h收取生菜叶用于后续试验或-20℃保存。
1.6 从江香猪源干扰素在生菜中瞬时表达的检测用剪刀在1.5收获的生菜叶片上随机剪1 cm×1 cm大小的叶片3片。将剪好的叶片置于1.5 mL离心管的无菌水中洗涤,用滤纸吸干叶片表面水分。按GUS染色液试剂盒说明书染色后肉眼观察显蓝色的植株后挑取完整的叶片放入载玻片中,滴一滴蒸馏水,400倍显微镜下观察并照相,检测各干扰素的表达情况。
分别称取真空侵染的生菜叶片4 g置于研钵中,加入液氮研磨后按每0.1g叶片加入150 μL蛋白提取 缓 冲 液 (NaCl, 8.9 g; KCl, 0.2 g; Na2HPO4,1.44 g,KH2PO4,0.24 g;ddH2O 定容至 1 000 mL,4℃保存),冰浴研磨。每隔10 min研磨一次,30 min后,4℃,12 000 r/min离心10 min,收集上清液即为蛋白液。按照猪IFN-α、猪IFN-β ELISA试剂盒说明分别检测从江香猪源IFN-α2、IFN-α2β、IFN-β在生菜中的表达。
1.7 生菜中瞬时表达的从江香猪干扰素蛋白对PRRSV抑制作用的检测 将1.6提取的各蛋白液利用BCA蛋白浓度测定试剂盒测定其浓度后,分别用滤菌器过滤并稀释至1.5 mg/mL。设置6个组:IFN-α2+PRRSV; IFN-β+PRRSV; IFN-α2β+PRRSV;IFN-α2+IFN-β+PRRSV,PRRSV 阳性对照;VEC(空载体)+PRRSV作为对照组。向长满Marc145细胞的24孔细胞培养板中每孔分别加入各蛋白液150 μL(蛋白终浓度为0.75 mg/mL),37℃5%CO2培养24 h,弃掉蛋白液后各孔加入300 TCID50的PRRSV病毒液,37℃ 5%CO2培养60 min,吸出病毒液,每孔加入 500 μL含 5%FBS的 DMEM,37℃ 5%CO2培养36 h后收集细胞。TRIzol试剂提取细胞总RNA反转录为cDNA,采用本实验室前期建立的荧光定量PCR(qPCR)检测Marc145细胞中PRRSV N基因的拷贝数[12]。采用相对比较 Ct法(Qr=2-ΔΔCt)分析数据,采用Graphpad_Prism_6.04软件绘制差异图。
向长满Marc145细胞的96孔细胞培养板中依次加入100 μL 4倍倍比稀释(41~410的生菜中表达的各从江香猪源的IFN),每个样设3个重复。设置5个 组 : IFN-α2+PRRSV; IFN-β+PRRSV; IFN-α2β+PRRSV;PRRSV;VEC+PRRSV。以VEC+PRRSV为对照组。37℃5%CO2培养24 h,弃掉蛋白液后各孔加入100 TCID50的PRRSV病毒液,37℃5%CO2培养60 min,吸出病毒液,每孔加入100 μL含5%FBS的DMEM,37℃5%CO2培养72 h后观察各IFN抑制Marc145细胞病变(CPE)的结果。
2 结果
2.1 含从江香猪源各IFN植物表达载体的构建与鉴定结果 采用1.3设计的引物PCR扩增从江香猪源各IFN基因,结果显示,扩增获得的目的条带分别约为540 bp、560 bp、1 100 bp,均与预期结果一致(图1A)。
利用 PCR 扩增获得的 IFN-α2、IFN-β、IFN-α2β基因分别构建重组质粒pBI121-IFN-α2、pBI121-IFN-β、pBI121-IFN-α2β。PCR 鉴定各重组质粒,结果显示扩增的各目的条带均与预期结果一致(图1B)。各重组质粒的双酶切鉴定结果均与预期一致(图略)。测序结果显示PCR扩增获得了大小分别为546 bp、561 bp、1 108 bp的基因序列,与GenBank登录的参考猪源 IFN-α2、IFN-β、IFN-α2β (融合基因)基因序列的同源性均达到99%。表明正确构建了含从江香猪源IFN植物表达载体pBI121-IFN-α2、
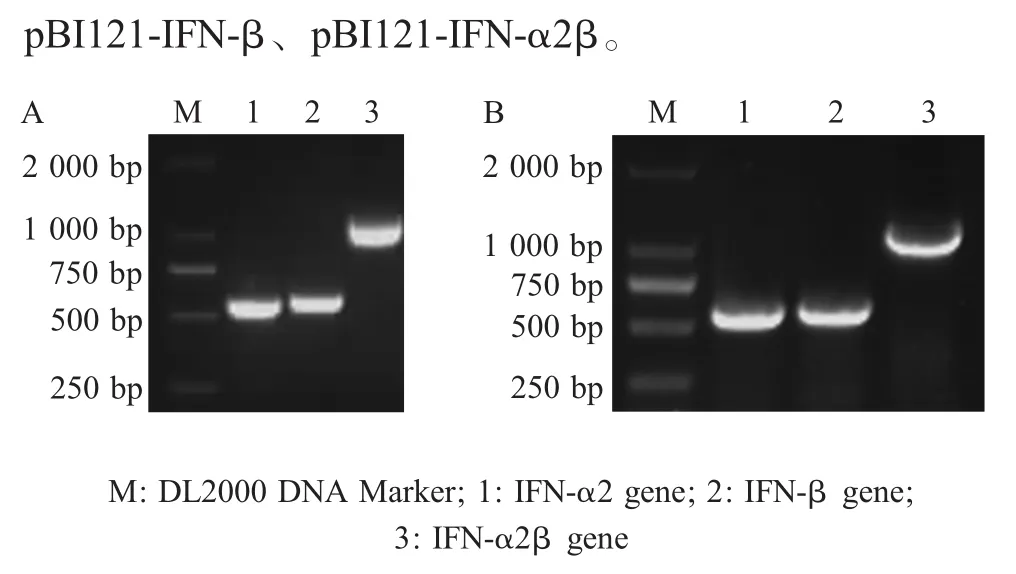
图1 IFN-α2、IFN-β、IFN-α2β 基因的 PCR 扩增(A)与各重组质粒的PCR鉴定(B)Fig.1 Amplification of IFN-α2,IFN-β,IFN-α2β genes(A)and identification of the each recombinant plasmids(B)by PCR
2.2 从江香猪源IFN在生菜中瞬时表达的检测结果含从江香猪IFN质粒的根癌农杆菌转染的生菜叶片经GUS染色。因PBI121载体融合外源基因后存在启动子而表达GUS基因;而空载体不存在启动子,则不表达GUS基因。在适宜条件下,若生菜表达各从江香猪IFN,那么也能够表达GUS。表达的GUS蛋白可以将GUS染液试剂盒中的X-Gluc水解生成蓝色产物。结果观察各干扰素质粒转染的生菜叶可见蓝色产物,而转染空载体的生菜叶未见蓝色产物(图2),初步表明,从江香猪源 IFN-α2、IFN-β、IFN-α2β蛋白在生菜叶中得到了瞬时表达。
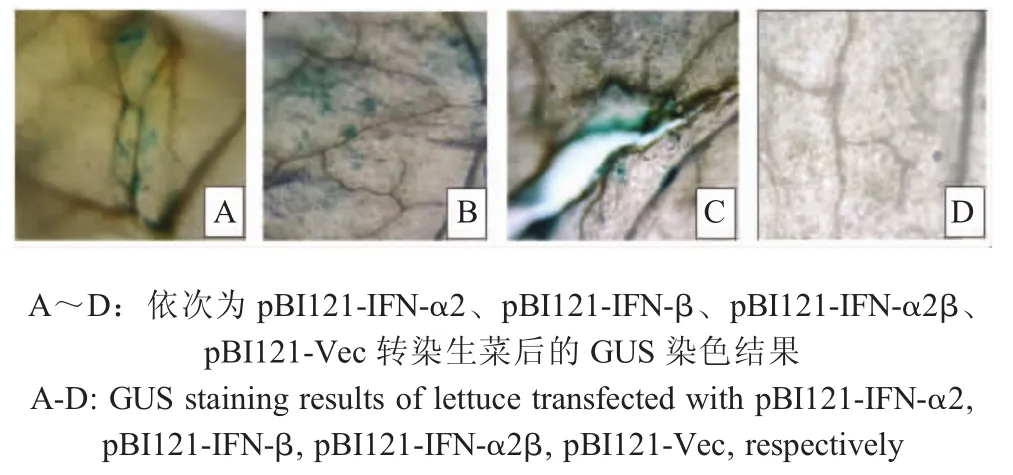
图2 从江香猪源IFN在生菜中表达的GUS得染色结果(10×40)Fig.2 GUS staining of lettuce transfected with Congjiangxiang porcine IFN(10×40)
2.3 从江香猪源干扰素生菜中瞬时表达蛋白的ELISA检测结果 利用BCA蛋白浓度测定试剂盒测定各IFN浓度,结果显示IFN-α2浓度为1.71 mg/mL;IFN-β浓度为1.80mg/mL;IFN-α2β浓度为3.12mg/mL;空载体(VEC)表达的总蛋白浓度为2.45 mg/mL。将各干扰素浓度稀释为1.50 mg/mL用于后续试验。
利用猪IFNα、IFN-β ELISA试剂盒检测从江猪源各 IFN 的含量。结果显示,IFN-α2、IFNα2β、IFN-β、IFN-α2β 蛋 白 含 量 均 值 分 别 为 3.25×10-8mg/mL、2.84×10-8mg/mL、1.51×10-7mg/mL、8.22×10-8mg/mL。进一步表明从江香猪源 IFN-α2、IFN-β、IFN-α2β在生菜中得到了瞬时表达,但其表达量偏低。
2.4 从江香猪源干扰素生菜中瞬时表达蛋白对PRRSV抑制作用检测结果 采用qPCR检测生菜中瞬时表达的从江香猪源 IFN-α2、IFN-β、IFN-α2β 对PRRSV复制的抑制作用,结果显示,从江香猪源IFN-α2、IFN-β蛋白对PRRSV复制具有显著抑制作用(p<0.01),融合表达的从江香猪源 IFN-α2β 蛋白抑制PRRSV复制的效果更显著(p<0.002)(图3)。将从江香猪源 IFN-α2、IFN-β、IFN-α2β 4 倍倍比稀释(41~410:3.75×10-1mg/mL~1.43×10-6mg/mL)处理Marc145细胞24 h,然后以100 TCID50PRRSV感染,观察结果显示,IFN-α2、IFN-β、IFN-α2β 分别经47、47、49稀释后仍然能够抑制 100 TCID50PRRSV的致CPE作用(表2)。表明融合表达 IFN-α2β蛋白比单一 IFN-α2或IFN-β对PRRSV复制的抑制效果更显著。
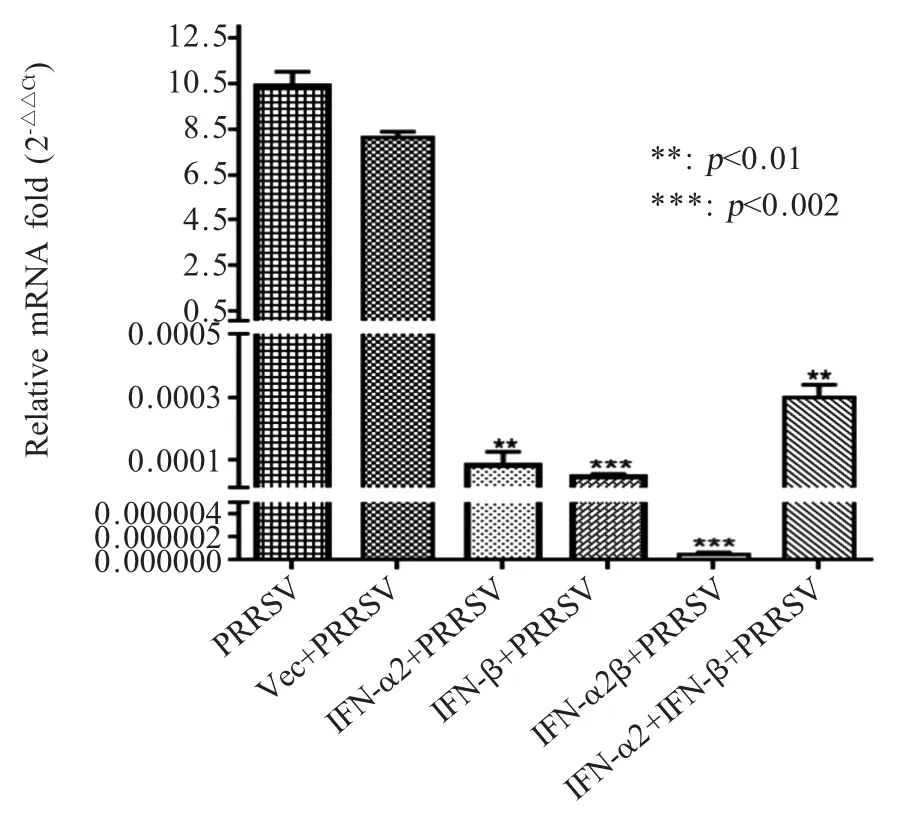
图3 生菜中瞬时表达的从江香猪源IFN对PRRSV复制抑制作用的检测结果Fig.3 Inhibition of PRRSV replication by transiently expressing IFNs from Congjiangxiang porcine in lettuce
表2 从江猪源IFN-α2、IFN-α2β、IFN-β 抑制 PRRSV的致CPE结果Table 2 The effect of congjiangxiang porcine IFN-α2,IFN-α2β,IFN-β on PRRSV induced CPE
3 讨论
猪IFN作为抗病毒先天免疫中关键性的细胞因子,主要通过增强免疫球蛋白的受体表达,激活免疫细胞,增强机体的免疫应答能力,从而干扰病毒的复制,属于新型抗病毒蛋白质类兽药[13]。郑鸣等通过大肠杆菌分别表达了猪IFN-α、猪IFN-β,均具有较好生物学活性[14];钟颖等[3]、刘敏等[4]、曹瑞兵[6]等用毕赤酵母表达猪IFN-α、猪IFN-β,表达产物也均具有较好的抗病毒活性。但以上各表达系统各有优缺点,大肠杆菌表达系统表达猪的IFN具有可操作性强、成本低等优点,但缺少真核细胞的细胞器,不能进行糖基化修饰及翻译。毕赤酵母表达系统虽具有大肠杆菌表达系统的良好可操作性和真核细胞的表达加工能力,但存在产量低及过度糖基化等问题[15]。因此有必要寻找一种新的经济、安全、快速、高产的猪源IFN表达体系。
植物表达体系具有高效、低廉、制备方法简单和规模化生产等独特的优势,ProdiGene公司用玉米表达了β-葡萄糖醛酸酶、胰蛋白酶和抗生物素蛋白;美国Ventria公司在水稻中表达了人乳铁蛋白、人溶菌酶等,均被批准作为精细化学产品投放市场[16]。生菜具有较高的营养价值,生长周期短,外源基因转入生菜中的转化效率较高,同时还可以避免药物蛋白在加热中的变性,是一种用来表达外源蛋白的良好宿主植物[17]。本研究中从江香猪源IFN在生菜中的表达量相对于罗熹等在生菜中表达的鸡IFN-γ[18]、李静在生菜中表达的人IFN-β的表达量偏低[19],推测可能受菌株限制,还有基因在生菜中的表达时间、蛋白提取方法、表达后检测时间等因素的影响,本实验室将开展采用不同菌株在生菜中表达从江香猪源的IFN,通过优化表达时间、蛋白提取方式、表达后检测时间等因素,以期获得更高的表达量,为从江香猪IFN蛋白的深入研究提供实验材料。
一个合理的动物源蛋白表达体系要实现价格低廉、安全、可控,而最主要问题就是保证植物表达蛋白的活性[20],以便用于治疗动物疾病。根癌农杆菌介导的瞬时转染方法已经被用来生产抗体分子[21]或糖基蛋白[20]。但是关于来源于该体系的动物源蛋白研究报道较少,本研究中证明了根癌农杆菌介导的从香江猪源IFN在生菜中的瞬时表达可以生产出高活性的IFN,同时融合从江猪源干扰素蛋白IFN-α2β具有天然从江猪源IFN的活性,对PRRSV的复制具有较强的抑制作用,显著优于IFN-α2或IFN-β或IFN-α2+IFN-β对 PRRSV复制的抑制作用,与郑鸣等在大肠杆菌中融合表达的猪IFN-α、IFN-β抗病毒活性结果一致[14]。该结果对抗PRRSV新型猪源IFN融合蛋白兽药的研制提供参考依据。同时根癌农杆菌介导的瞬时转染方法也可以作为未来生产猪源融合蛋白的一种有效方式。
