盐碱胁迫对碱地肤的生物量及含氮化合物的影响
2019-09-02田浩然刘航铄何刘贵杰
田浩然,杨 傲,刘航铄,何刘贵杰,赵 航,麻 莹
(吉林大学植物科学学院,吉林 长春 130062)
我国东北草原盐碱化已经超过70%[1]。通常盐碱化土壤既含有中性盐又含有碱性盐,中性盐胁迫和碱性盐胁迫实际是两种不同性质的胁迫[2],分别称为盐胁迫和碱胁迫。盐胁迫和碱胁迫的作用原理及植物对其所作出的生理响应是不同的。然而,从研究现状来看,仍以盐胁迫为主,目前只有少数报道涉及到了碱胁迫[3-5]。
氮素是植物体内最重要的营养元素,氮代谢则是其体内最重要的物质代谢。通过影响氮素的吸收、同化、无机氮到有机氮的转化,以及蛋白质、核酸等含氮生物分子的合成等生理过程,氮代谢与植物的生长、光合等基本生命活动密切相关[6]。在研究植物抗逆生理机制时,氮代谢的研究也是不可忽视的。例如,在镉[7]、砷[8]等重金属胁迫以及高温[9]、干旱[10]、盐[11]等非生物胁迫下,均发现植物含氮化合物及其相关酶活性发生了显著变化。
碱地肤(Kochia sieversianus)是一种高度抗碱的盐生牧草(可以在pH > 10的土壤中生存),其植株黄绿色或稍带红色,上端密被白色绒毛。碱地肤还具有一定的药用价值,其全草及种子均可以作为中药材使用[1]。由于其既具有极强的耐盐碱能力又具有很好经济价值,而成为中国东北地区治理碱化草地的首选牧草之一。以碱地肤为材料研究植物抗盐碱生理机制、比较抗盐、抗碱机制的异同,无疑是一种理想的选择。而有关盐或碱胁迫对碱地肤含氮化合物的影响差异目前尚不了解。为此,将碱性盐(Na2CO3和NaHCO3)和中性盐(NaCl和Na2SO4)分别以等摩尔比混合,模拟出碱胁迫和盐胁迫条件以胁迫处理碱地肤幼苗,通过监测其生物量及各种含氮化合物含量随胁迫时间的动态变化,来比较盐、碱胁迫对碱地肤氮素积累影响的差异,以期为研究碱地肤的耐盐碱机制提供必要的理论依据,同时也为进一步探讨高抗盐碱植物碱地肤特殊的氮代谢调节机制奠定理论基础。
1 材料与方法
1.1 幼苗培养
从吉林省西部天然草原采集碱地肤种子,用砂培法培养[4]。出苗后每天17:00用1/2 Hoagland营养液浇灌。每盆16株幼苗(花盆内直径20 cm)。幼苗培养、处理均在室外进行,处理时期人工避雨。
1.2 试验设计
将Na2SO4和NaCl两种中性盐以1∶1摩尔比混合作为盐胁迫组(A),将碱性盐Na2CO3和NaHCO3以同样比例混合作为碱胁迫组(B)。盐胁迫组和碱胁迫组的总盐浓度都是200 mmol·L-1。这样可保证盐胁迫组和碱胁迫组处理液的Na+、总离子浓度相同,而只有两组的pH不同[4]。以Hoagland营养液作为对照组(CK)。盐胁迫组(A)、碱胁迫组(B)和对照组(CK)处理液的pH分别为6.58、9.96及6.56[4]。苗龄6周后,将66盆碱地肤(选取长势一致的幼苗)随机分成22组,每组3盆即3次重复[4]。其中1组在处理开始时即取样测定起始生物量及各种含氮化合物含量,另外7组为对照组(CK),对照组只浇Hoagland营养液在7个时间点(16,24,36,48,72,96,144 h),此外,盐胁迫处理和碱胁迫处理各7组(盐、碱胁迫处理时间点与对照组相同)。含有相应浓度胁迫盐的营养液为处理液,每盆500 mL处理液透灌[4]。
1.3 取样及指标测定
取样:按照对应的处理时间,依次取出全部植株[4]。分别用自来水和蒸馏水清洗植株,之后吸干植株表面水分。将植株分成茎叶和根两部分(以子叶痕处为界),记录各组茎叶部分的鲜重(fresh weight,FW)。另外从以上样品中称取10 g茎叶部分冻干,记录其干重。将冻干样品粉碎混匀,用于硝态氮(NO3--N)、氨态氮(NH4+-N)、脯氨酸、甜菜碱等含量测定;其余鲜样杀青(105 ℃,15 min)后置于真空干燥箱内(40 ℃)干燥至恒重,计算总干重(dry weight,DW)。指标测定:生物量 = 地上干重(DW)/株。含水量 = (FW-DW)/FW × 100%。硝态氮(NO3--N)用离子色谱法测定[1](美国戴安DX-300离子色谱系统,AS4A-SC离子交换柱,流动相为Na2CO3/NaHCO3=1.7/1.8 mmol·L-1, CDM-II电 导 检 测 器 )。 氨 态 氮(NH4+-N)采用改良的茚三酮测定法[12]。脯氨酸和甜菜碱分别用酸性茚三酮法[12]和紫外分光光度法[12]进行测定。
1.4 数据分析
采用SPSS 19.0统计分析;数据采用平均值 ±标准误表示,检验水平为0.05;Excel 2010制图。
2 结果
2.1 盐、碱胁迫对碱地肤幼苗生物量的影响
非胁迫(对照组)和盐胁迫条件下,碱地肤地上干重随着时间的延长呈现显著增加的趋势(P< 0.01),而且对照组生物量的增加幅度(F= 25.43)高于盐胁迫(F= 5.62)。碱胁迫条件下,其生物量增加并不明显(P> 0.05) (图1)。可见,盐或碱胁迫条件,特别是碱胁迫不利于碱地肤生物量的积累。
盐或碱胁迫条件下,碱地肤地上含水量均呈现降低趋势(P< 0.01)。而碱胁迫的(F= 191.80)下降幅度显著高于盐胁迫(F= 29.57),但是在长时间(144 h)的高碱胁迫(pH ≈ 10)下碱地肤保持着较高的含水量 (≈ 75%) (图 1)。
2.2 盐碱胁迫下无机氮的动态积累特点
在非胁迫条件下(对照组),NO3--N、NH4+-N含量并未随时间变化而变化(P> 0.05)。NO3--N含量在盐、碱胁迫16 h时出现峰值,之后其含量均急剧下降,36 h后盐胁迫下含量保持不变(P> 0.05),而碱胁迫下含量有所升高(图2)。在盐、碱胁迫条件下NH4+-N含量先升高而后降低,而且碱胁迫处理下NH4+-N的积累始终高于盐胁迫(图2)。此外,盐、碱胁迫促NH4+-N积累的最高峰出现的时间点也不同。盐胁迫处理NH4+-N最大积累量出现在48 h,而碱胁迫的时间在72 h。
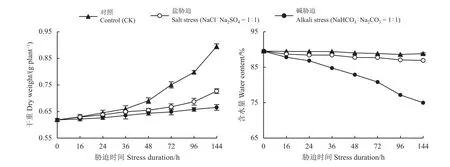
图1 盐碱胁迫下生物量及其含水量的动态变化Figure 1 Time-course of changes in biomass and water content under salt and alkali stress
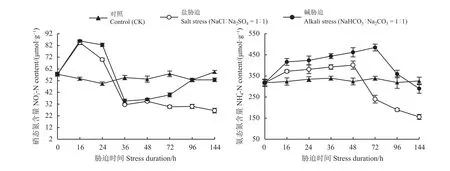
图2 盐碱胁迫下无机氮含量的动态变化Figure 2 Time-course of changes in inorganic nitrogen contents under salt and alkali stress
2.3 盐碱胁迫下有机氮的动态积累特点
在盐、碱胁迫下,甜菜碱和脯氨酸均随着胁迫处理时间的延长而有所积累,其中碱胁迫的作用大于盐胁迫(图3)。脯氨酸的变化趋势是在碱胁迫处理48 h内增加缓慢,一旦超过48 h,脯氨酸含量显著增加(P< 0.01),且其含量的最高峰出现在96 h;盐胁迫下脯氨酸含量随时间延长而增加,而且在盐处理72 h时脯氨酸达到积累最大量,之后有所降低。甜菜碱含量的转折点出现在盐碱胁迫24 h处,之后其含量均有所升高,而且在碱胁迫下其含量高于盐胁迫。
3 讨论
3.1 盐、碱胁迫对碱地肤生物量的影响
生长受到抑制是植物对盐碱胁迫普遍而直接的反应[13],生长抑制的直接表现就是生物量降低。与对照组(非胁迫条件)相比,盐、碱胁迫均抑制碱地肤生物量的积累,其中碱胁迫对其抑制作用更强。碱地肤为适应碱胁迫(高pH)需要耗费更多的能量和物质,因此碱胁迫对碱地肤生物量的抑制更为严重。这是在逆境下碱地肤重要的生存策略之一。
降低体内含水量是植物响应渗透胁迫进行渗透调节的一种既迅速又经济的方式[1]。在本研究中,随着盐、碱胁迫处理时间的延长,碱地肤茎叶的含水量随之下降,而碱胁迫的(F= 191.80)下降幅度显著高于盐胁迫(F= 29.57)。此外,与抗盐碱能力强的禾本科植物芦苇(Phragmites australis)[14]、狗牙根(Cynodon dactylon)[15]以及药用盐生植物宁夏枸杞(Lycium barbarum)[15]不同,碱地肤在长时间(144 h)的碱胁迫条件下仍能保持较高的含水量(75%),高含水量可以最大限度地浓缩细胞质中溶质浓度,稀释液泡中有害离子的浓度[14.16]。而碱地肤并不是肉质化植物,它在长时间的离子毒害和高pH条件下还保持着较高的含水量,这也许就是决定碱地肤植物既抗盐又抗碱的关键生理特性之一,这一结果值得深入研究。
3.2 盐、碱胁迫对碱地肤含氮化合物的影响
生物量的变化是抗盐碱植物碱地肤适应盐碱环境的外在表现,而碱地肤体内含氮化合物的变化才是响应盐碱胁迫的内在机制。氮是植物生长发育不可或缺的营养元素。植物中氮主要以有机氮和无机氮两种形式存在。无机氮的形式主要以硝态氮(NO3--N)和氨态氮(NH4+-N)存在,二者也是植物吸收氮素的主要形式[17]。研究表明,当植物遭受盐或碱胁迫等逆境条件时,NO3-在维持渗透平衡和离子稳态中起着重要作用[18]。NO3--N在盐、碱处理144 h内先后出现“升高-下降-平缓”的趋势,但碱胁迫下其含量始终高于盐胁迫,此结果与甜土植物向日葵(Helianthus annuus)、大麦(Hordeum vulgare)不同,盐、碱胁迫均抑制向日葵、大麦茎叶NO3--N积累,且碱胁迫抑制作用更大[19-20]。可见,作为抗盐碱植物碱地肤,其特殊的NO3--N吸收机制还有待我们继续研究。
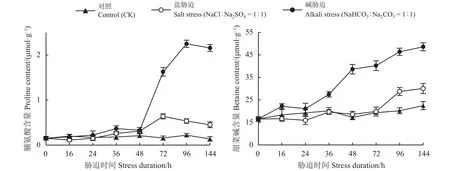
图3 盐碱胁迫下有机氮含量的动态变化Figure 3 Time-course of changes in organic nitrogen contents under salt and alkali stress
植物体内NO3-可被逐步还原为氨态氮(NH4+-N),之后与其自身吸收的NH4+-N一起经过一系列生理生化反应最终转化为有机氮形式,被植物体利用[21]。用GEOCHEM软件分析盐、碱胁迫处理液中离子活度和自由度表明,与盐胁迫相比,碱胁迫处理液中大部分NH4+-N转化为NH3[22]。在碱胁迫处理液中NH4+-N大量减少的情况下,本研究中虽然盐、碱胁迫下NH4+-N的含量均呈现先升高而后下降的趋势,但碱胁迫下NH4+-N的含量始终高于盐胁迫(图2)。这可能与碱胁迫下体内大量NO--3N还原为NH4+-N及氮素的分配等方面有关,具体原因需要进一步研究。此外,盐胁迫(48 h)、碱胁迫(72 h)促进NH4+-N积累的最高峰出现的时间点也不同。综上,盐、碱胁迫对碱地肤NH4+-N的积累存在差异。
盐生植物在遭受非生物胁迫时,通常会合成一些以含氮化合物为主的相容性物质,如季胺类化合物,脯氨酸和甜菜碱等[21]。盐胁迫下植物脯氨酸的大量积累常作为衡量植物耐盐性的标志[23]。而本研究结果表明,脯氨酸在碱胁迫48 h后才积累,并且其含量极低(其含量占总有机溶质的0.3%~1.1%)。可见,脯氨酸完全失去了一个敏感胁变指标的特点。它的积累可能不是响应渗透胁迫的结果,而可能是一种抗盐碱的适应性防御反应。此外,在植物细胞生理干旱条件下,脯氨酸的增加还有利于保持细胞或组织的持水能力,提高植物在盐渍胁迫的适应性。
研究表明,甜菜碱主要在叶绿体中合成,甜菜碱在植物体内合成后几乎不被分解[24],本研究中,对照组(CK)的甜菜碱含量(占干重 2.4%)很高,碱地肤体内甜菜碱含量高可能与其抗盐碱逆境的调节机制有关。随着盐碱胁迫时间的延长,甜菜碱含量持续增加(最高可占干重3.5%),而且在碱胁迫下其含量高于盐胁迫。
4 结论
与盐胁迫相比,碱胁迫严重抑制碱地肤的生长(生物量、含水量降低),更促进含氮化合物(脯氨酸、甜菜碱)的积累。可见,碱胁迫对碱地肤氮代谢的影响更为严重,碱地肤响应碱胁迫的氮代谢调节机制是非常复杂的,值得深入探讨。也再次印证了盐、碱胁迫是两种不同性质的胁迫,碱胁迫对植物的影响更复杂、更严重[2]。
