两种栽培条件下柳枝稷的分株构件生物量分配与生长分析
2019-09-02胡宗好杨允菲李海燕
胡宗好,杨允菲,李海燕
(东北师范大学草地科学研究所 / 植被生态科学教育部重点实验室,吉林 长春 130024)
植物的生物量分配特征是植物自身和环境因素共同作用的结果,它反映出植物对环境的适应能力和生长发育规律[1]。其中叶是植物重要的物质生产器官[2],而繁殖是物种延续和种群发展的前提,因此叶生物量分配和生殖分配在植物的生长繁殖过程中占居着重要的地位。在复杂的环境中,植物通过某些性状,比如个体大小、分蘖数量以及生物量分配等,调节策略以保证顺利完成种群的延续[3]。植物除了在不同的环境中具有不同的生长和分配策略,它在生长发育过程中,自身生理状况会发生改变,其生长和分配策略也会不断调整,但前提是遵循生长与物质分配相协调原则[4]。
柳枝稷(Panicum virgatum)为禾本科(Gramineae)黍属(Panicum),多年生C4草本植物,起源于北美,植株高大、根系发达,具有较强的耐旱耐贫瘠能力,对环境的适应能力很强[5-7]。柳枝稷可以作为牧草和能源原料,其细胞壁可被消化为糖类并随之发酵生产燃料乙醇,生产成本低,很有发展潜力[8]。以往有关柳枝稷的研究主要集中在光合生态生理[9-11]、种子萌发和幼苗生长[12-14]、干旱胁迫和盐胁迫生理[15-17]等,关于柳枝稷的生物量分配和生长分析的研究迄今尚很少报道[18]。
在松嫩平原上,不论是沙壤土还是盐碱土,均可以栽种柳枝稷,但不同的土壤条件,其生长和生产具有较大的可塑性,这种可塑性则蕴涵着生长与物质分配策略[4]。本研究对沙壤土和盐碱土栽培的柳枝稷种群,于乳熟期和蜡熟期进行两次大样本分株取样调查,就其分株的数量特征进行了土壤类型间的对比分析,以及有关生物量分配的相关性分析,以期揭示柳枝稷的生长和物质分配策略,为不同生长条件下柳枝稷的生物学与生态学特性的深入研究提供科学积累和理论参考。
1 材料与方法
1.1 样地的自然概况
试验地设在松嫩平原南部吉林省松原市长岭县种马场东北师范大学草地生态研究站内(44°38′ N,123°41′ E)。该区属于温带半湿润、半干旱季风型气候[19]。
样地1设在东北师范大学生态研究站的5 hm2实验田内,柳枝稷种植面积为10 m × 100 m,土壤为沙壤土(sandy soil,S);样地2设在盐碱化草甸的开垦栽培实验区内,柳枝稷种植面积为8 m × 100 m,土壤为盐碱土(saline-alkali soil,Sa)。
1.2 取样与测定
2017年9月上旬(乳熟期)和10月初(蜡熟期),对2016年春季沙地实验区和盐碱地实验区播种的二年生柳枝稷分株分别做大样本取样,在每种土壤栽培试验区随机抽取30株,齐地面剪下,分别测定各个分株高度,编号分置。在恒温箱80 ℃烘干至恒重,使用1/1 000分析天平分别称量叶片生物量、叶鞘生物量、茎生物量和花序生物量。
1.3 数据处理
对于测定的分株数量和生物量,用IBM SPSS Statistics 24.0对其进行独立样本t检验,并计算各项数量指标的平均数(mean)、标准差(SD)和变异系数(CV);采用Excel 2013对分株性状间的关系进行相关性分析与t检验并制图,在线性、指数和幂函数3种函数中,取相关程度最高的函数作为定量分析模型;采用花序生物量占分株生物量的百分比作为生殖分配的数量指标。其计算公式为:
RA(生殖分配) = (花序生物量/分株生物量) × 100%。
2 结果与分析
2.1 分株的数量特征
经统计分析,柳枝稷分株数量特征在乳熟期和蜡熟期,均为沙壤土极显著大于盐碱土(P< 0.01)(表1)。其中生物量上(叶片生物量、叶鞘生物量、花序生物量、茎生物量、分株总生物量),沙壤土乳熟期各项均值分别为盐碱土的2.08、2.08、3.32、2.13和2.39倍;蜡熟期分别为1.62、1.60、2.74、1.85和1.97倍。SPSS分析表明,在变异系数上,各项指标在种群内的变异系数均小于种群间,即种群内的变异度小于种群间。
对比柳枝稷在乳熟期和蜡熟期的分株数量特征,柳枝稷分株的各项指标,蜡熟期相对于乳熟期的均值均有所提升。
2.2 分株数量特征之间的关系
分析表明,两种土壤类型上栽培的柳枝稷分株株高与分株总生物量具有不同的规律性变化(表2)。沙壤土上种植的柳枝稷在乳熟期株高和分株总生
物量呈线性同速增长规律,而到了蜡熟期则呈现为幂函数异速增长规律。盐碱土上种植的柳枝稷在乳熟期为幂函数异速增长,到了蜡熟期则变成线性同速增长。

表1 两种栽培条件下的柳枝稷种群分株的数量特征及其显著性检验Table 1 Quantitative characteristics of ramets and significance test of switchgrass populations under two planting conditions
株高与分株构件生物量的关系因土壤条件和生育期的不同而异。在沙壤土上,乳熟期,株高与叶片生物量、茎和叶鞘生物量均呈指数相关关系;蜡熟期,呈幂函数关系。在盐碱土上,乳熟期,叶片生物量与株高呈线性相关关系,茎+叶鞘生物量呈指数相关关系;在蜡熟期,株高与叶片生物量呈幂函数关系,与茎+叶鞘生物量则呈线性关系。
分株生物量和分株构件生物量也因土壤条件和生育期的不同而异。在沙壤土上,两个生育期的分株生物量与叶片生物量均呈幂函数相关关系;在盐碱土上,乳熟期,分株生物量与叶片生物量呈线性相关关系,蜡熟期则呈幂函数相关关系。但两种土壤条件下,茎和叶鞘生物量在两个生育期均呈线性关系。由此反映了不同土壤类型栽培的柳枝稷在不同生育期其分株的生长与各构件生物量分配上存在一定的调节,致使规律发生了变化。
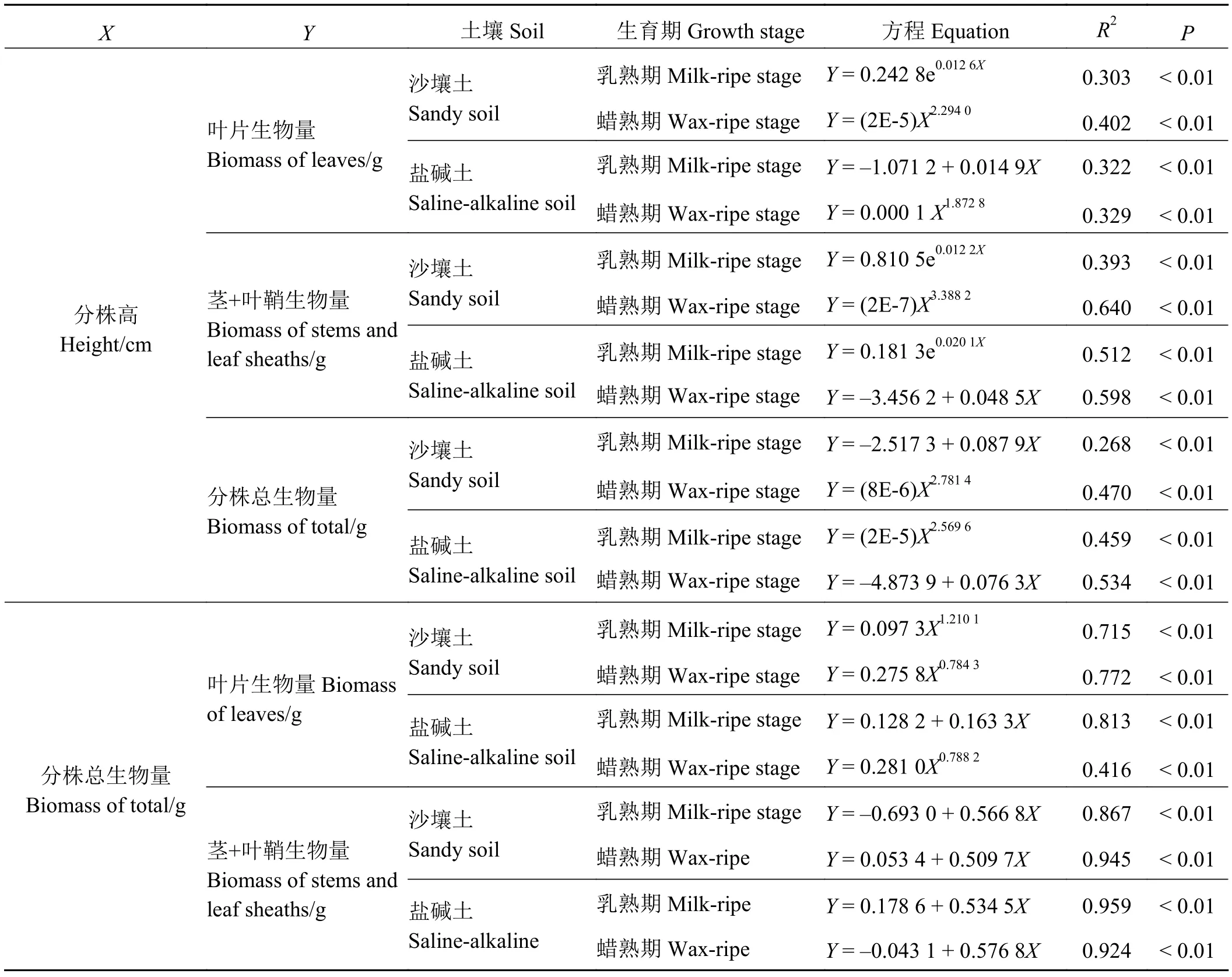
表2 两种栽培条件下的柳枝稷种群分株数量特征的定量关系及其显著性检验Table 2 Quantitative relationships of ramets and significance test in switchgrass populations under two planting conditions
2.3 分株生物量分配
统计发现,两种土壤分株生物量分配均以茎比重最大(表3)。在乳熟期和蜡熟期两个生育期,分株茎比重在盐碱土分别为42.90%和41.94%,大于沙壤土的37.40%和39.05%;均以叶鞘的生物量分配比重最小,盐碱土分别为14.63%和15.03%,大于沙壤土的12.76%和12.40%;而生殖分配则表现为沙壤土(33.45%和31.59%)大于盐碱土(23.14%和22.63%)。两种土壤类型之间,只有叶片生物量分配在蜡熟期差异不显著(P> 0.05),除此之外,叶鞘、茎和花序生物量分配均达到显著差异(P< 0.05)。对同一土壤类型不同生育期之间的统计分析表明,柳枝稷分株各构件生物量分配均没有显著差异。由此反映了在乳熟期以后,两种土壤柳枝稷分株生物量分配格局尚未发生大幅度调节。
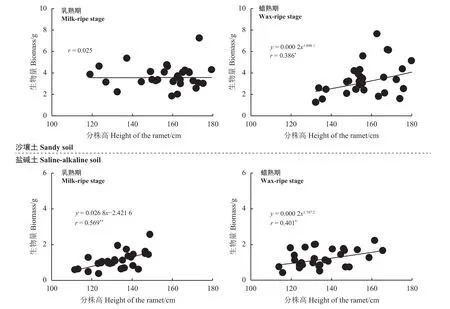
图1 两种栽培条件下的柳枝稷种群花序生物量与株高的关系Figure 1 Relationships between inflorescence biomass and ramet height of switchgrass populations under two planting conditions
2.4 花序生物量和生殖分配与株高之间的关系
相关性分析和显著性检验(图1)表明,柳枝稷花序生物量与分株高之间,在沙壤土的乳熟期无显著相关性(P> 0.05),到了蜡熟期则呈显著的幂函数正相关关系(P< 0.05);在盐碱土的乳熟期花序生物量与分株高之间呈极显著线性正相关关系,到了蜡熟期则呈显著幂函数正相关关系。由此反映出,不同土壤类型栽培的柳枝稷在不同生育期其分株越高,其花序生物量也越大,并且存在一定的调节而使其规律发生了变化。
在生殖分配与分株株高之间,在沙壤土的乳熟期表现为极显著的指数负相关关系,蜡熟期则呈显著的线性负相关关系;在盐碱土的乳熟期和蜡熟期均不具有显著相关性(图2)。由此反映出,沙壤土种植的柳枝稷在不同生育期其分株越高,其生殖分配也越小,其规律也发生了变化;而盐碱土生殖分配至乳熟期以后一直相对恒定。

图2 两种栽培条件下的柳枝稷种群生殖分配与株高的关系Figure 2 Relationships between reproductive allocation and ramet height of switchgrass populations under two planting conditions
3 讨论与结论
在松嫩平原,9月上旬与10月初分别是柳枝稷生长的乳熟期期和蜡熟期,同一样地内的柳枝稷生长数量特征在两段时间内无显著差异,这说明柳枝稷在乳熟期到蜡熟期,其各项特征在生物量和形状上均变化不大。在两个样地间,沙壤土上的柳枝稷在各项数量特征大于盐碱土上的柳枝稷,说明,相对于盐碱土而言,柳枝稷更适合生长于沙壤土。两个时期,两种土壤间分株的多数特征变异系数均大于土壤内分株的数量特征的变异系数,并且通过方差分析,土壤间柳枝稷数量特征差异显著,说明在这些特征上土壤间的柳枝稷生态可塑性大于土壤内。
不同土壤上栽培的柳枝稷,分株的株高和分株生物量之间的生长关系上也表现出不同的生长规律。同一土壤类型上栽培的柳枝稷在不同时期也表现出不同的生长规律,总的来说,在蜡熟期较乳熟期其生长规律更加稳定。沙壤土上柳枝稷在乳熟期表现为线性同速增长,到了腊熟期,沙壤土柳枝稷以幂函数趋势异速增长,这表明,在腊熟期分株生物量随着株高变化加大。沙壤土柳枝稷分蘖数量较多,株丛较为茂盛密集,在生长过程中分株占据的空间极为重要,株高对光资源的影响也越来越大。相反,盐碱土柳枝稷在乳熟期呈幂函数趋势异速增长,意味着在恶劣环境的旺盛生长期,较大植株具有较强的生长优势,到了腊熟期表现为线性同速增长,这可能与盐碱土上的柳枝稷分蘖数量较少、株丛较为稀疏有关,即在没有强烈光资源竞争的情况下,相对较矮的分株得到了充分生长,致使其分株生物量变化随着株高增加趋于平缓。
分株株高的增加和有机物质的增重都属于生长的范畴[20]。生长期不同、土壤环境不同,分株的组分生物量同株高的函数变化规律也会发生变化。如果把这种函数规律看作是遗传因子控制的,用拟合方程的确定系数R2来估计,随机环境影响用1-R2来估计。在分株株高同叶片生物量、茎叶生物量的关系中分别有30.27%~53.42%、39.33%~64.02%是由遗传控制的。而分株生物量同叶片生物量、茎叶生物量的关系中分别有41.64%~81.32%、86.66%~95.94%是由遗传控制的。并且在不同时期、不同土壤其R2也有较大的差异,说明柳枝稷在不同的土壤中和不同时段,其生长和物质分配均具有较大的生态可塑性和可调节性。
在生物量分配上,两个时期,不同土壤上栽培的柳枝稷均表现为茎 > 花序 > 叶 > 叶鞘,就功能而言,植物的叶片是营养物质的产生器官,茎具有养分输导和储藏以及支撑叶片扩展生长空间的多重作用[4]。这两种土壤上栽培的柳枝稷在种群的水平上表现为,优先分配到茎,然后是花序,最后是叶和叶鞘。说明柳枝稷群落中,分株把较多的营养物质用于茎的伸长生长,以提高对空间和光资源的竞争力;然后是花序,也就是意味着柳枝稷将较多的能量用于繁殖上。但是,全部样本生物量分配的变异系数在10.9%~30.3%,意味着样本内总是存在或大或小的变化,如果把样本的各个数量特征作为一个连续的变化过程,则对于生殖分配和株高之间却呈负相关关系,表明分株的高生长要以减少其生殖分配为代价[21-22],这也体现了植物生长的一种调节策略[23-25]。对比这两种土壤上的柳枝稷来看,沙壤土上种植的柳枝稷叶、茎、叶鞘的生物量分配低于盐碱土上种植的柳枝稷,而生殖分配高于盐碱土上种植的柳枝稷,可能与沙壤土的生长环境优于盐碱土有关。因为盐碱土上种植的柳枝稷分株株丛较为稀疏,种内竞争较小,但植株需要将更多的能量用于抵御不良环境[26],而恶劣环境对大小分株的影响是一致的,这也是盐碱土柳枝稷生殖分配一直处于恒定状态的原因。
植物在生长发育中,在遵循生长和物质分配相协调的原则上,不断地调整着植物生殖生长的生物量分配策略[27]。在沙壤土环境下,乳熟期柳枝稷分株花序生物量表现的较为恒定,到了蜡熟期花序生物量与株高之间呈现出正相关关系,但生殖分配一直随着株高的增加而减少;在盐碱土环境下,株高和花序生物量一直保持着正相关关系,而生殖分配却表现的较为恒定。这反映出,柳枝稷具有较大的生态可塑性以及较强的可调节性,在应对不同环境时,用不同的生长策略以保证自身的生长发育和繁殖。
