短期高温胁迫对海岛棉不同部位果枝产量形成的影响*
2019-08-31张巨松阿不都卡地尔库尔班郭仁松
陈 振, 陈 平, 张巨松**, 阿不都卡地尔·库尔班, 林 涛, 郭仁松
短期高温胁迫对海岛棉不同部位果枝产量形成的影响*
陈 振1, 陈 平3, 张巨松1**, 阿不都卡地尔·库尔班1, 林 涛2, 郭仁松2
(1. 新疆农业大学农学院/教育部棉花工程研究中心 乌鲁木齐 830052; 2. 新疆农业科学院经济作物研究所 乌鲁木齐 830091; 3. 阿瓦提县气象局 阿瓦提 843200)
高温是影响棉花产量的重要环境因子, 研究盛花期短期高温胁迫对海岛棉不同部位果枝产量及产量构成因素的影响, 有助于为海岛棉选育耐热稳产性品种、制定丰产抗逆栽培技术提供理论方法。以‘新海43号’和‘新海49号’为材料, 于盛花期在田间搭设增温棚, 设置不同增温持续时间[0 d(CK)、3 d(H3)、6 d(H6)、9 d(H9)]模拟短期高温胁迫对海岛棉干物质积累、叶片净光合速率、蕾铃脱落率、产量及产量构成因素的影响。结果表明: 盛花期短期高温胁迫, 导致‘新海43号’和‘新海49号’中部果枝蕾铃脱落率显著增加, 棉株总干物质积累量下降。在花后30~50 d, 各增温处理棉铃的干物质量与对照相比显著降低; 而花后40~50 d, 两品种H6、H9处理茎、叶干物质量与对照相比显著升高。随生育进程的推进, ‘新海43号’和‘新海49号’在H3、H6、H9处理下棉铃最大干物质量比对照分别降低8.9%、29.3%、36.3%和11.8%、28.1%、42.6%; 棉铃日均积累量分别下降10.9%、32.8%、42.2%和12.8%、30.3%、45.9%; 而达到快速积累期终止时期所需的时间分别增加5 d、8 d、14 d和1 d、5 d、10 d。在增温处理期间, 两品种主茎叶n随增温持续时间的延长呈大幅下降趋势; 增温结束后, 各处理叶片n的衰减速度不同, 在生殖生长后期(花后40 d) H3、H6、H9处理下叶片n均高于对照, 表现为H9>H6>H3>CK。短期高温胁迫导致两品种单株结铃数、单铃重和衣分显著降低, 其中单株结铃数变异系数(15.4%~18.5%)最大, 衣分变异系数(4.1%~4.7%)最小。‘新海43号’和‘新海49号’H3、H6、H9处理下单株结铃数分别减少21.6%、22.5%、28.9%和16.6%、26.4%、34.7%; 而增温显著增加了上部果枝铃数, 进而提高了上部果枝产量及产量贡献率。品种间, ‘新海49号’净光合速率、单株结铃数和单铃重下降幅度以及蕾铃脱落率的增加幅度均大于‘新海43号’, 说明‘新海43号’具有更好的耐热性。
海岛棉; 高温胁迫; 果枝位置; 干物质积累; 净光合速率; 产量
高温是一种重要的非生物胁迫, 严重限制了许多地区作物的生长与发育[1]。IPCC第5次评估报告中指出, 在过去30年间, 全球表面平均温度升高了0.85 ℃。随着全球表面温度的上升, 大部分陆地发生高温极端事件的频率将增高, 持续时间将会更长[2]。在近50年中, 中国年平均地表气温升高1.1 ℃, 中国三大棉区增温显著且增温面积不断扩大[3]。因此, 温度的升高是影响当前棉花(spp.)生产的重要因素之一。
海岛棉()是一种重要的经济作物, 是纺织高档和特种织物的重要原料, 其优良的品质在世界范围内享有盛誉, 目前其产量只占世界棉花产量的2%左右[4-5]。新疆是中国唯一的海岛棉生产区, 新疆海岛棉常年种植面积为10万hm2, 总产量约占世界海岛棉的1/4, 在我国具有重要的经济与战略地位[6]。棉花的生长过程不仅受自身遗传的控制, 还受到外界环境因子的影响。高温胁迫对棉花的生理、生化特性都产生严重的影响, 进而影响棉花的生长发育及产量品质[7-8]。随着全球气候变暖, 温度的上升必将对我国海岛棉的生长发育产生重要影响。
棉花对高温非常敏感[9]。先前的研究表明, 对温度较为敏感的生育期内, 温度每升高1 ℃棉花产量将减少17%[10]。而棉花处于高温胁迫下, 日最高温度每升高1 ℃, 将可能导致皮棉产量减少110 kg∙hm-2[8]。花铃期对高温更为敏感, 过高的气温会使花粉生活力下降, 从而导致大量蕾铃脱落[11]。根据源库理论, 产量的形成是光合作用与干物质转移共同决定的[12]。Oosterhuis[13]研究发现, 高温导致棉花产量下降的根本原因是碳水化合物的供应不足, 碳水化合物供应不足主要是因为棉株“源”端受到高温的影响。同时Loka等[14]也进行了类似的研究, 夜间增温导致棉花呼吸速率显著增加, 叶片ATP水平和碳水化合物含量降低, 最终导致产量下降。
以往的研究多集中于长期高温对棉花生殖生长的影响[15-16]。然而, 随着极端高温胁迫事件发生的频率越来越高, 短期高温胁迫对棉花的影响越来越受到关注。余新隆等[17]认为, 在棉花盛花期连续遭受3 d以上高温, 会造成花粉粒畸形, 花药不开裂, 影响花粉的正常授粉受精。Shi等[18]研究发现, 短期的高温胁迫会对水稻()后期的光合作用与物质转移产生持续的影响。由于棉花的生长具有不同步性, 在不同的果枝部位、不同的时间、不同的环境条件下形成了具有不同发育速度的棉铃[19]。因此, 短期的高温胁迫, 必将导致不同果枝棉铃对高温反应的差异。以往的研究主要集中在高温对整株陆地棉产量的影响上, 目前鲜有关于高温胁迫对海岛棉不同部位棉铃影响的研究。本文以海岛棉作为受试材料, 在大田条件下, 研究了短期高温胁迫对海岛棉干物质积累、净光合速率及不同部位果枝产量和产量构成因素的影响, 为进一步开展气候变暖条件下海岛棉的响应与适应研究, 以及制定海岛棉抗逆栽培技术提供理论依据。
1 材料与方法
1.1 试验设计
试验于2018年在新疆阿瓦提县长绒棉研究开发中心试验基地进行。试验地土质为砂壤土, 0~60 cm耕层土壤pH 7.8, 有机质含量为5.7 g∙kg-1, 全氮为0.6 g∙kg-1, 水解性氮为50.3 mg∙kg-1, 速效磷为 19.6 mg∙kg-1, 速效钾108 mg∙kg-1。海岛棉全生育期每月平均温度与降雨量情况如图1所示。
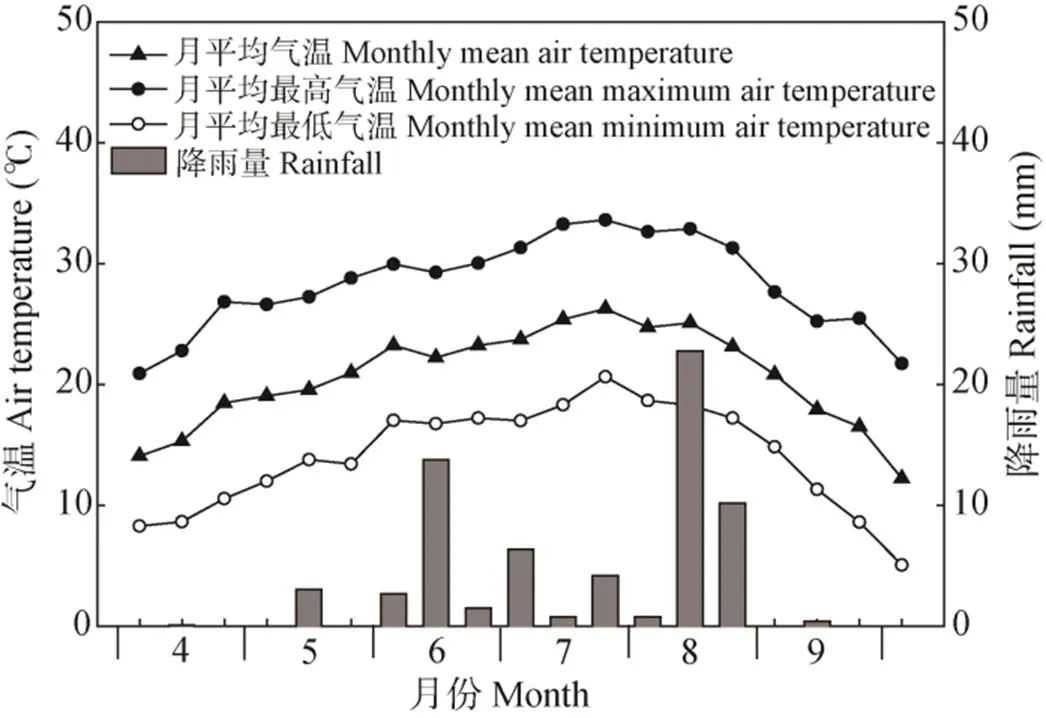
图1 2018年研究区海岛棉全生育期月气温(平均、最高和最低)与降雨量情况
供试材料为‘新海43号’和‘新海49号’两个海岛棉品种。前者在近年异常高温天气下种植, 具有较好的结铃率, 产量未受明显影响; 后者具有较好的产量品质性状, 表现出很强的生态适应性, 目前在南疆大面积种植。试验采用二因素(品种、增温持续时间)随机区组设计, 3次重复, 小区面积为32.5 m2。两品种的盛花期开始增温处理: 以大田自然状态为对照(CK); 结合南疆地区近37年高温天气的实际情况, 设置3 d(H3)、6 d(H6)、9 d(H9) 3个增温处理, 即各处理连续3 d、6 d、9 d增温。‘新海43号’高温处理时间为7月5—15日; ‘新海49号’高温处理时间为7月13—23日。增温棚采用焊制的铁管骨架为构架, 塑料薄膜厚度为0.08 mm, 透光率为91%, 高1.5 m, 面积与小区面积相同。在增温处理期间, 每天11:00—20:00盖棚模拟高温环境。棚的四周留有距地面30 cm高度间隙, 以允许充分的气体交换和避免过度增温。温湿度记录仪(GM1365, 深圳市标智科技有限公司)挂在棚内外距冠层上方30 cm处, 每隔1 h自动记录小区温度, 处理与对照田的温度如图2所示。处理结束后拆除增温棚, 恢复到与对照一致的环境水平直至成熟。除以上高温处理外, 海岛棉整个生育期的肥水和病虫害管理均按大田进行。
1.2 测定项目及方法
1.2.1 干物质测定
于海岛棉花后(第6果枝50%开花, 下同)0 d、10 d、20 d、30 d、40 d和50 d天取样, 每小区取代表性植株5株, 按茎、叶、棉铃分样, 105 ℃下杀青30 min, 85 ℃烘干至恒重后称重。
1.2.2 主茎功能叶净光合速率(n)测定
主茎功能叶光合作用用TPS-2便携式光合仪(英国PP Systems公司)测定系统测定。于花后0 d、3 d、6 d、9 d、20 d、30 d、40 d和50 d选择晴朗天气于11:30—13:30测定主茎功能叶净光合速率(n)。每处理每重复取3片生长一致且受光方向相近的主茎功能叶测定。
1.2.3 产量及产量构成因素测定
按不同的果枝节位(FB)把棉株分为3部分: 下部果枝(第1~5节果枝, FB1-5)、中部果枝(第6~10节果枝, FB6-10)和上部果枝(第11及以上节果枝, FB≥11)。在收获期, 每小区定30株进行单株单铃收获, 准确记录收获铃所在果枝节位, 统计单株铃数及各部位铃数, 单铃称重, 计算平均单铃重。然后将棉铃按上、中、下3部分分别轧花, 测定衣分。并计算蕾铃脱落率:
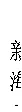
1.3 数据处理
试验数据使用SPSS 23.0软件进行方差分析, 采用Duncan法进行处理间多重比较(<0.05), 利用origin 8.5整理数据并绘图。
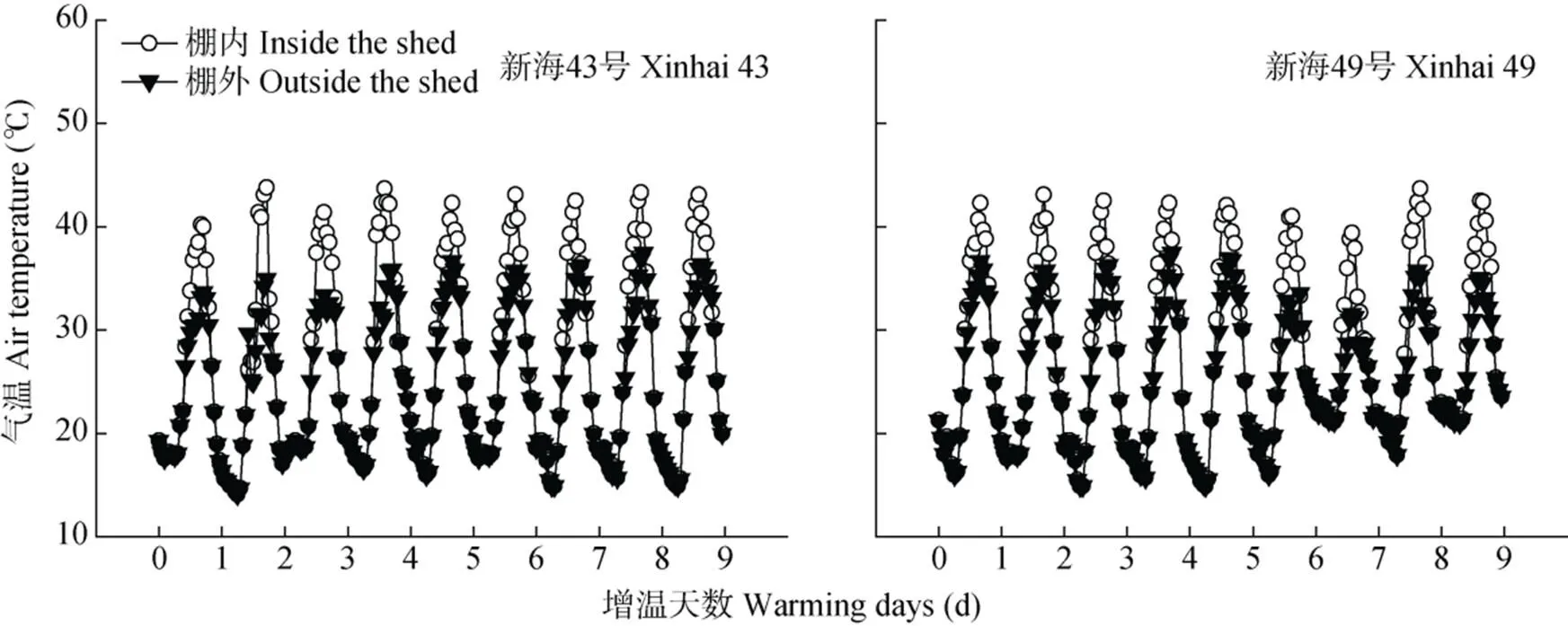
图2 2018年海岛棉盛花期高温胁迫期间高温处理(棚内)和对照(棚外)的温度变化
2 结果与分析
2.1 短期增温对海岛棉干物质量的影响
如图3所示, 随增温天数增加, 植株总干物质积累量下降, 在花后30~50 d, 各处理棉铃的干物质量与对照相比显著降低(<0.05), 两品种表现出相同的特点, ‘新海43号’H3、H6、H9处理棉铃干物质量分别比对照减少13.1%~20.4%、34.0%~35.1%、43.2%~52.6%; ‘新海49号’H3、H6、H9处理棉铃干物质量分别比对照减少13.1%~15.1%、29.9%~ 33.2%、49.4%~51.6。在花后40~50 d, 两品种H6、H9处理茎、叶干物质量与对照相比显著升高(<0.05), ‘新海43号’H6、H9处理茎叶干物质量分别比对照增加16.0%~16.4%、16.4%~23.2%; ‘新海49号’H6、H9处理茎叶干物质量分别比对照增加11.1%~19.0%、13.2%~23.6%。表明茎、叶器官在生殖生长后期储存了大量的营养物质。
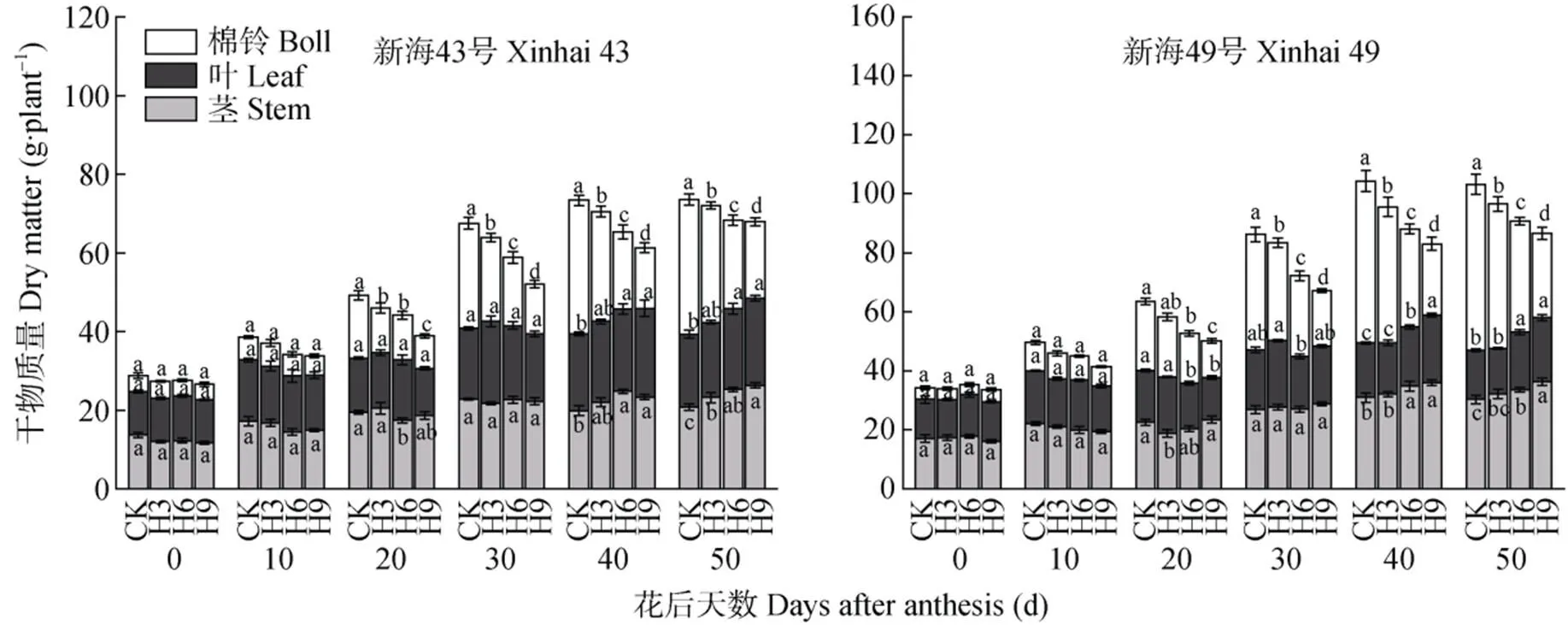
图3 盛花期短期增温下不同海岛棉品种地上部各器官干物质量的动态变化
CK: 不增温; H3: 增温3 d; H6: 增温6 d; H9: 增温9 d。不同小写字母表示同一品种不同处理间在0.05水平差异显著。CK: no warming; H3: warming for 3 days; H6: warming for 6 days; H9: warming for 9 days. Different lowercase letters indicate significant differences among treatments of the same organ at 0.05 level.
用Logistic生长函数对各处理棉铃干物质积累进行拟合, 其拟合度均达显著水平(<0.05)。由表1可知, 随生育进程的推进, 棉铃的最终干物质积累量降低, 最大积累速率减小, 到达快速积累期终止时期所需的时间延长。与对照相比, ‘新海43号’和‘新海49号’在H3、H6、H9处理下棉铃最大干物质量分别降低8.9%、29.3%、36.3%和11.8%、28.1%、42.6%; 棉铃日均积累量分别下降10.9%、32.8%、42.2%和12.8%、30.3%、45.9%; 而达到快速积累期终止时期所需的时间分别增加5 d、8 d、14 d和1 d、5 d、10 d。表明短期增温推迟了棉铃的发育。
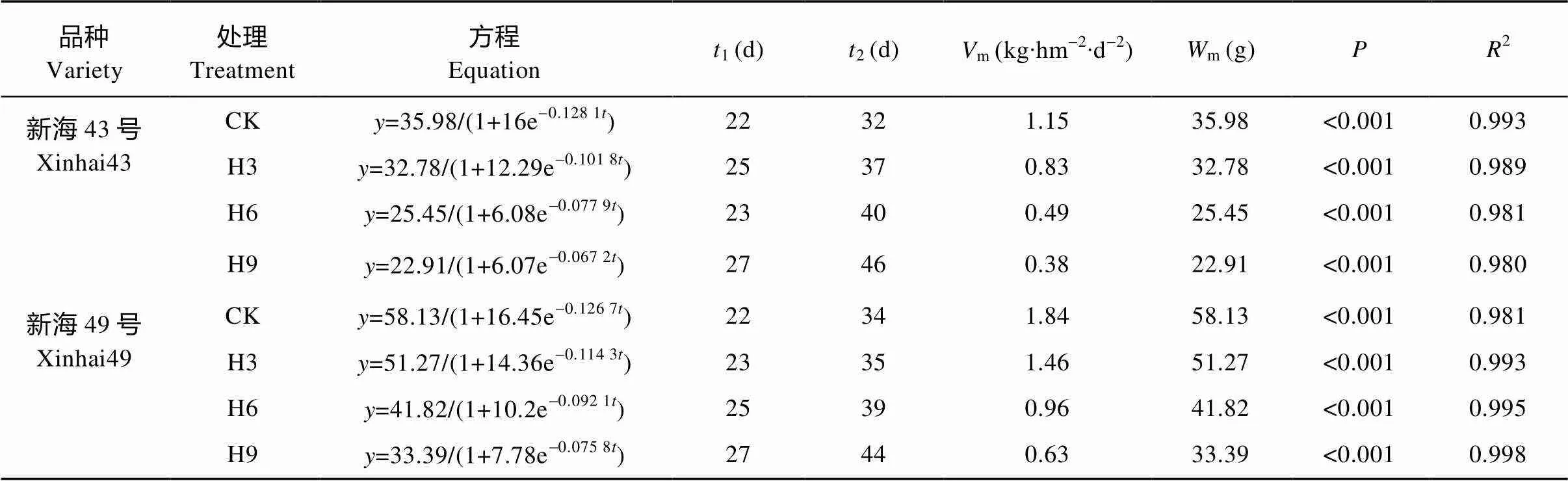
表1 盛花期短期增温下海岛棉棉铃干物质积累的Logistic模型及其特征值
CK: 不增温; H3: 增温3 d; H6: 增温6 d; H9: 增温9 d。: 棉铃干物质累积;: 花后天数(d);1: 最大积累速率时间;2: 快速积累期终止时间;m: 棉铃干物质最大累积速率;m: 最大理论干物质量;: 回归概率值。CK: no warming; H3: warming for 3 days; H6: warming for 6 days; H9: warming for 9 days.: boll dry matter accumulation;: days after anthesis;1: maximum accumulation rate time;2: terminating date of fleet accumulation period;m: maximum dry matter accumulation rate of boll;m: maximal theoretical biomass accumulation value;: probability of regression.
2.2 短期增温对海岛棉净光合速率(Pn)的影响
由图4可知, 盛花期增温结束后, 海岛棉主茎叶n在外界环境中随生育时期的推进而降低, 但各处理间的差异在花后不同生育阶段不一致。在增温处理期间, ‘新海43号’和‘新海49号’主茎叶n随增温持续时间的延长呈大幅下降趋势; 增温结束后,各处理叶片n的衰减速度不同。在生殖生长后期(花后40 d), H3、H6、H9处理下的植株n均高于对照, 但H3处理与对照并无太大差异。花后50 d, 两品种的n总体表现为H9>H6>H3>CK。表明短期增温在较短时间内对叶片伤害程度较大,n下降显著, 但随生育期推进会得到一定程度的修复。

图4 盛花期短期增温对不同海岛棉品种净光合速率(Pn)的影响
CK: 不增温; H3: 增温3 d; H6: 增温6 d; H9: 增温9 d。CK: no warming; H3: warming for 3 days; H6: warming for 6 days; H9: warming for 9 days.
2.3 短期增温对海岛棉产量及产量构成因素的影响
由表2可知, 在产量构成因素中, 增温对海岛棉的单株结铃数、单铃重、衣分和籽棉产量都具有显著影响(<0.05)。变异系数(CV)结果表明, 单株结铃数对增温最敏感, 衣分对增温最不敏感。在H3、H6、H9处理下, ‘新海43号’和‘新海49号’的籽棉产量较对照分别下降11.4%、20.5%、27.3%和12.0%、31.4%、44.4%, 而籽棉产量的降低主要是由于单株结铃数分别减少了21.6%、22.5%、28.9%和16.6%、26.4%、34.7%。
由表3可知, 增温与果枝部位对单株结铃数、籽棉产量的影响存在显著交互效应(<0.05); 品种与果枝部位对单株结铃数、单铃重和籽棉产量的影响存在显著交互效应(<0.05)。而品种、增温水平和果枝部位, 以及品种和增温水平的交互对单株结铃数、单铃重、衣分和籽棉产量均未达到显著水平(>0.05)。
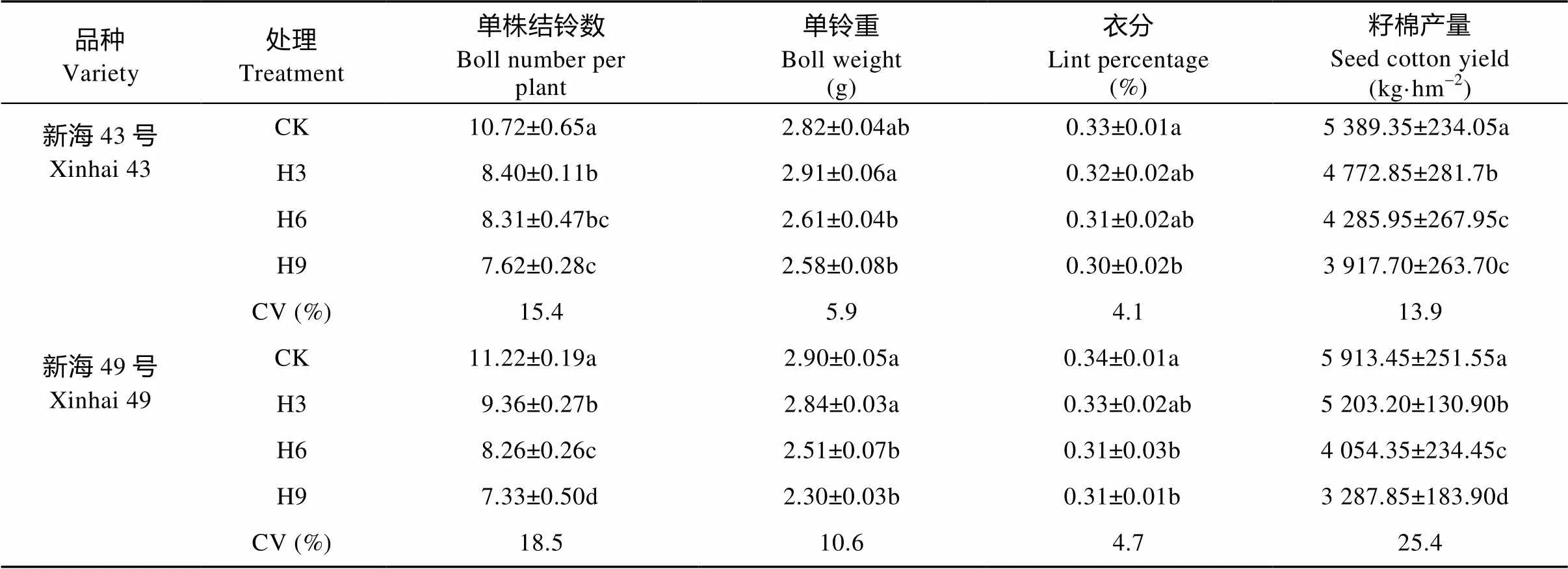
表2 盛花期短期增温对海岛棉产量及产量构成因素的影响
CK: 不增温; H3: 增温3 d; H6: 增温6 d; H9: 增温9 d。CV%: 变异系数。不同小写字母表示同一品种不同处理间在0.05水平差异显著。CK: no warming; H3: warming for 3 days; H6: warming for 6 days; H9: warming for 9 days. CV: coefficient of variation. Different lowercase letters indicate significant differences among treatments of the same variety at 0.05 level.
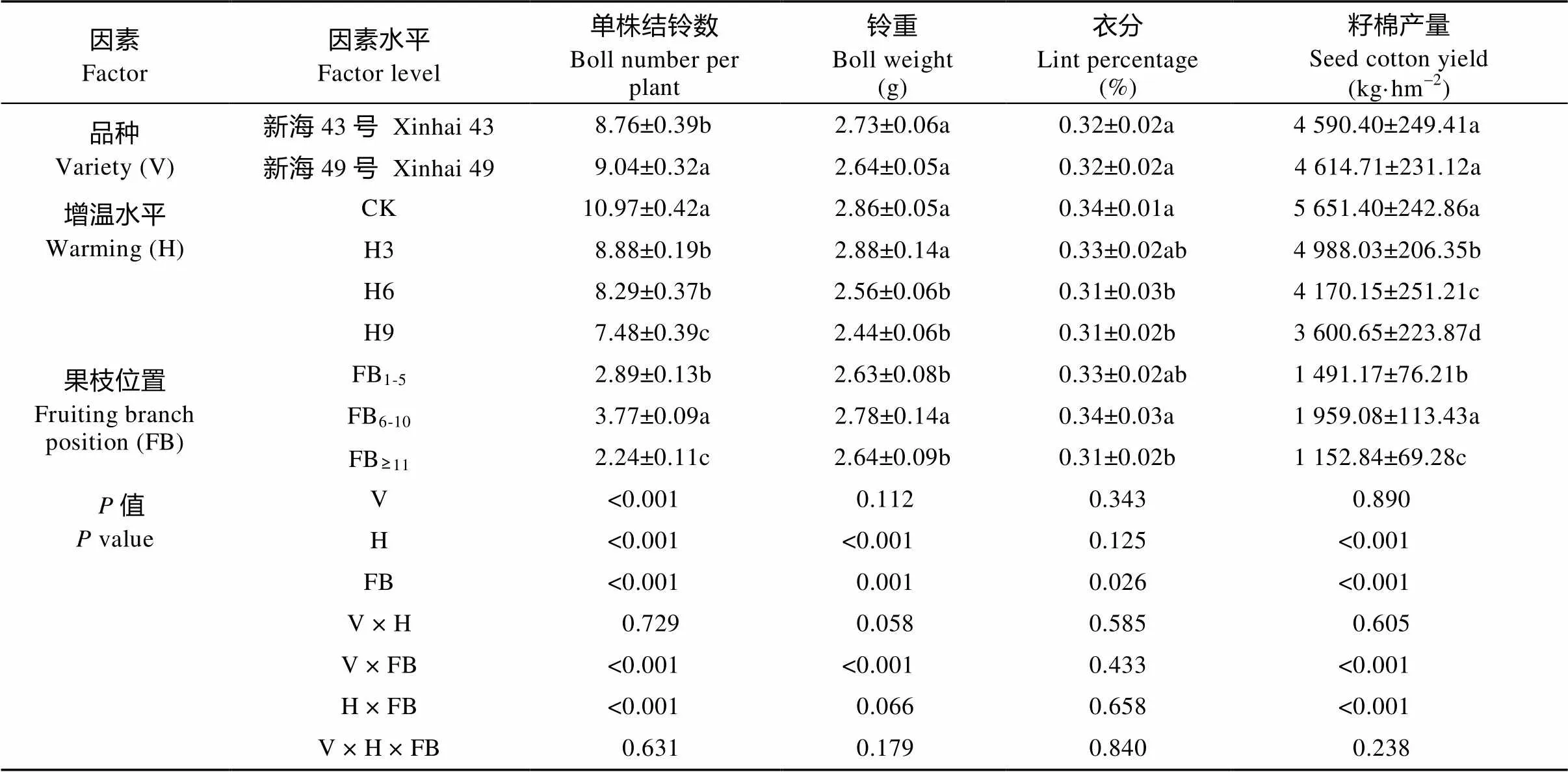
表3 盛花期增温、品种和果枝位置在产量及产量构成因素上的统计学意义及交互作用
CK: 不增温; H3: 增温3 d; H6: 增温6 d; H9: 增温9 d。FB1-5: 下部果枝; FB6-10: 中部果枝; FB≥11: 上部果枝。不同小写字母表示同一因素不同水平间在0.05水平差异显著。CK: no warming; H3: warming for 3 days; H6: warming for 6 days; H9: warming for 9 days. FB1-5: lower fruiting branches; FB6-10: middle fruiting branches; FB≥11: upper fruiting branches. Different lowercase letters indicate significant differences among different levels of the same factor at 0.05 level.
进一步分析不同部位果枝产量及产量构成因素的差异发现(表4、表5), 随着增温天数的延长, 下部和中部果枝的铃数、单铃重、衣分、籽棉产量逐渐下降, 蕾铃脱落率则呈逐渐上升的趋势; 其中中部果枝均出现显著性差异(<0.05), 而增温对下部果枝影响不大。与此相反, 增温条件下, ‘新海43号’和‘新海49号’上部果枝籽棉产量较对照分别增加0~18.7%和5.9%~28%; 两品种在H6、H9处理下上部果枝的单铃重、衣分显著降低(<0.05), 而铃数分别增加20%~24.9%和31%~58.2%, 且增温水平及增温水平与品种的交互作用对上部果枝铃数的影响均达显著水平(<0.05)。此外, ‘新海43号’和‘新海49号’ H3 处理上部果枝皮棉贡献率与对照相比无显著差异, 而两品种H6、H9 处理分别比对照显著增加49.2%~50.7%和65.5%~130.2%。
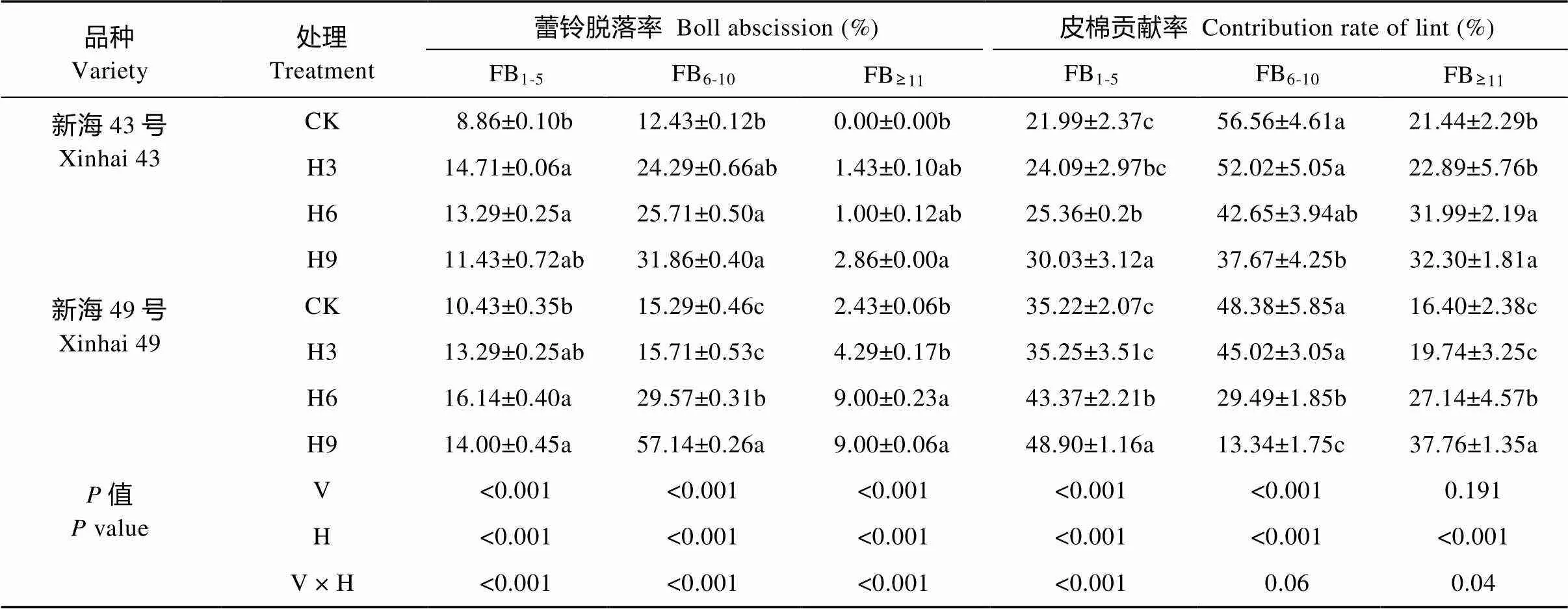
表4 盛花期短期增温对海岛棉不同部位果枝蕾铃脱落率及皮棉贡献率的影响
CK: 不增温; H3: 增温3 d; H6: 增温6 d; H9: 增温9 d。FB1-5: 下部果枝; FB6-10: 中部果枝; FB≥11: 上部果枝。V: 品种; H: 增温。同列不同小写字母表示同一品种不同处理间在0.05水平差异显著。CK: no warming; H3: warming for 3 days; H6: warming for 6 days; H9: warming for 9 days. FB1-5: lower fruiting branches; FB6-10: middle fruiting branches; FB≥11: upper fruiting branches. V: variety; H: warming during full-blooming stage. Different lowercase letters in the same column indicate significant differences among treatments of the same variety at 0.05 level.
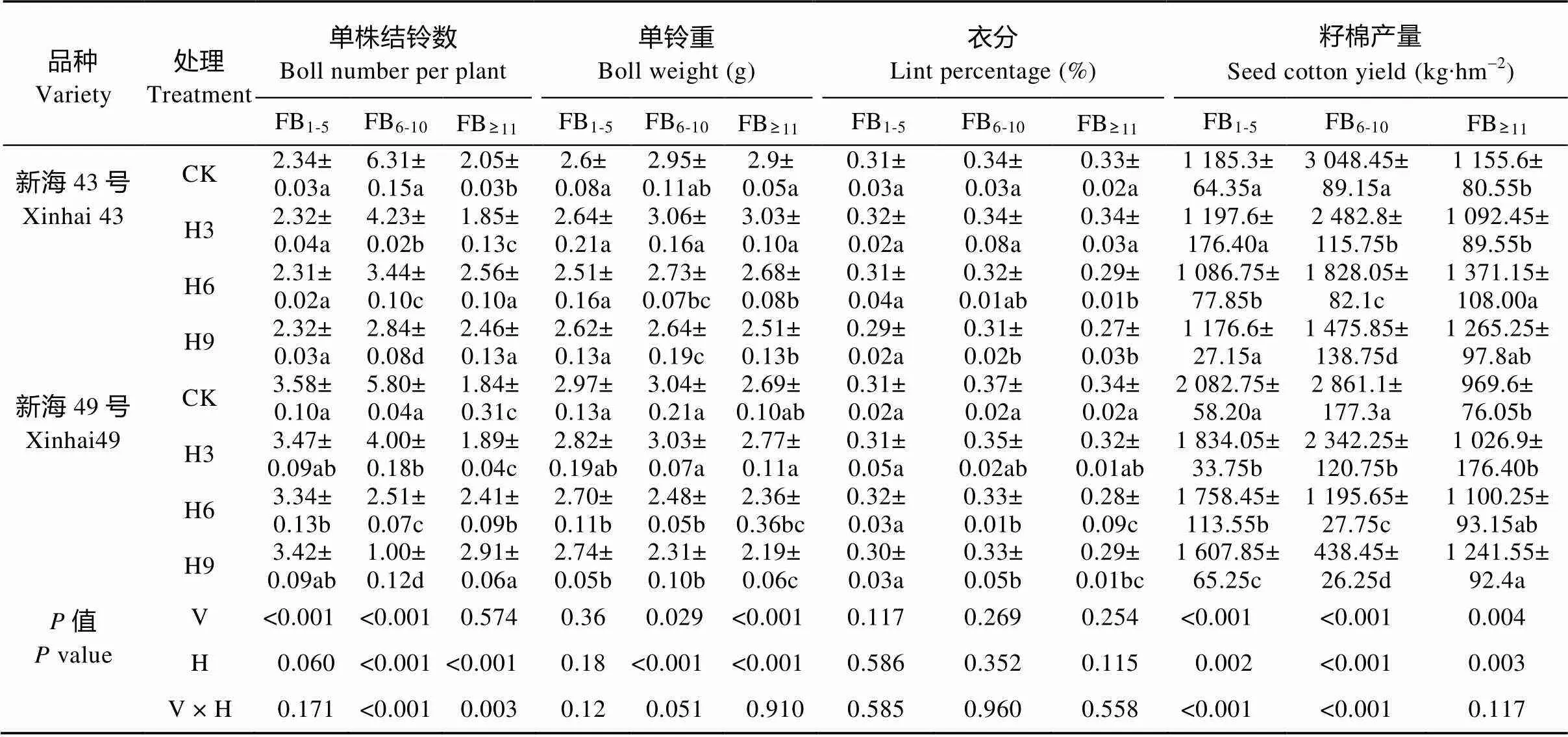
表5 盛花期短期增温对海岛棉不同部位果枝产量及产量构成因素的影响
CK: 不增温; H3: 增温3 d; H6: 增温6 d; H9: 增温9 d。FB1-5: 下部果枝; FB6-10: 中部果枝; FB≥11: 上部果枝。V: 品种; H: 增温。同列不同小写字母表示同一品种不同处理间在0.05水平差异显著。CK: no warming; H3: warming for 3 days; H6: warming for 6 days; H9: warming for 9 days. FB1-5: lower fruiting branches; FB6-10: middle fruiting branches; FB≥11: upper fruiting branches. V: variety; H: warming. Different lowercase letters in the same column indicate significant differences among treatments of the same variety at 0.05 level.
盛花期短期增温对海岛棉各部位果枝造成的影响存在差异。图5分别量化了短期增温对两品种各部位果枝产量的影响。结果表明, 增温时间每持续1 d, ‘新海43号’和‘新海49号’的下部果枝产量分别减少13.22 kg∙hm-2和43.13 kg∙hm-2, 中部果枝分别减产173.73 kg∙hm-2和269.55 kg∙hm-2, 上部果枝产量分别增加18.68 kg∙hm-2和29.53 kg∙hm-2。总体来看, ‘新海43号’产量的下降幅度较‘新海49号’小, 表明增温对‘新海49号’产量影响更大。
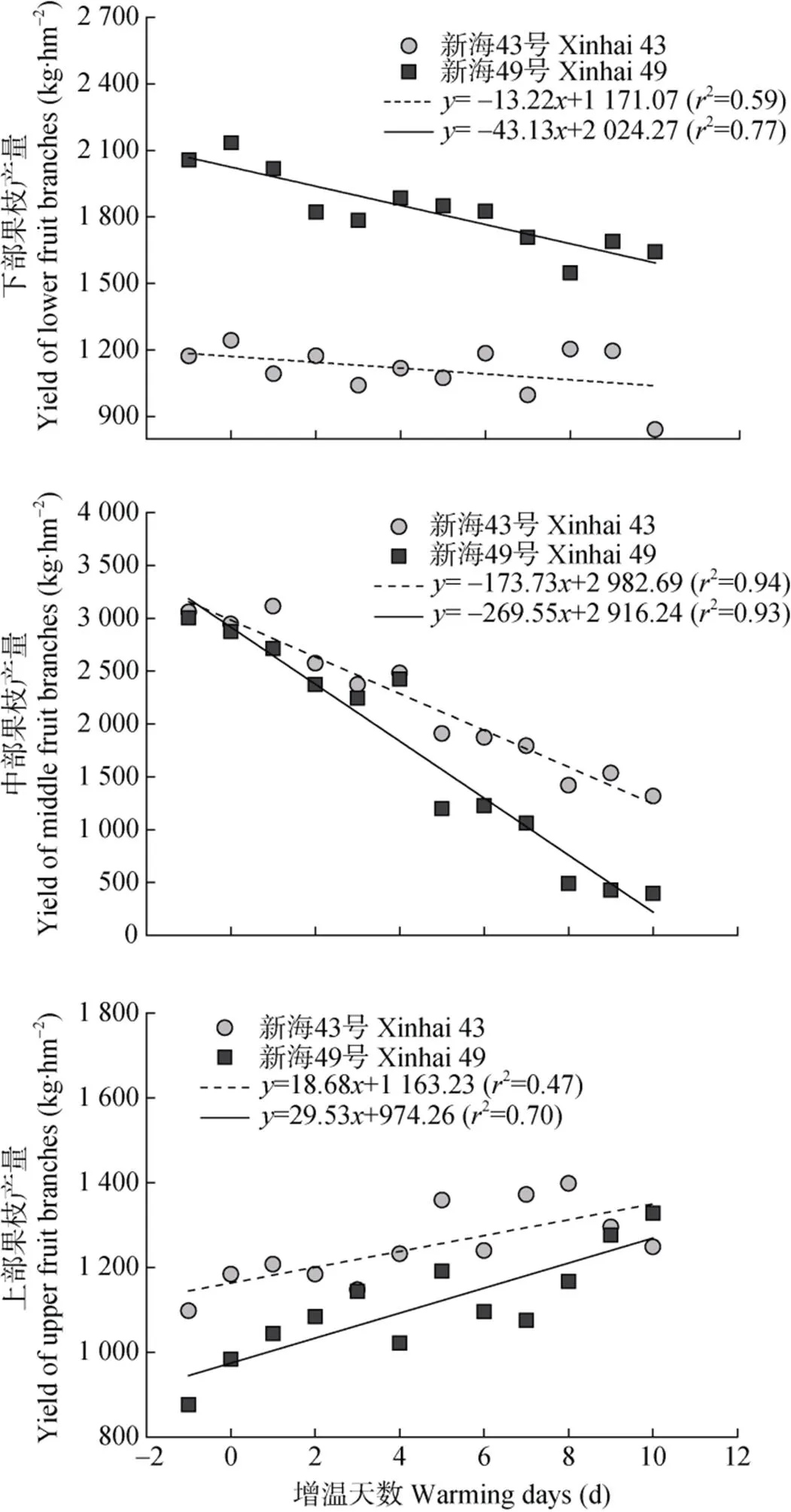
图5 盛花期短期增温下不同海岛棉品种不同部位果枝产量与增温天数之间的关系
3 讨论
在以往的大多数研究中, 棉花产量及其构成因素都是在整株上测定, 从而平均了外部胁迫作用对更高果枝棉铃的影响[20-21]。Brook等[22]研究表明, 棉花产量与补偿效应之间存在很强的交互作用。这一发现也许解释了为什么在全株水平上计算出来的单铃重和产量之间没有显著性的差异[23]。
作物高产是以较高的生物量为前提, 而高温极易影响棉株“源”端光合产物的储存和转运, 导致碳水化合物向库端的供应减少, 进而显著影响棉花的产量[13,24]。贺新颖等[7]认为, 花铃期增温2~3 ℃下, 棉株干物质积累总量下降10%左右, 主要表现为生殖器官生物量显著降低, 而营养器官生物量则基本不变。Singh等[8]研究发现, 高温胁迫导致蕾铃脱落率升高、单铃重降低以及畸形铃增多。本研究条件下, 棉株总生物积累量随增温时间的延长呈下降趋势, 其中主要是由于棉铃生物量显著降低; 而生殖生长后期棉株的茎、叶生物量却显著上升, 这可能与增温导致中部蕾铃大量脱落, 源库比严重失调, 叶片光合产物无法继续向棉铃器官输送, 导致部分光合产物在营养器官中堆积有关。
不同部位果枝棉铃的发育具有不同步性, 这可能导致对增温不同的反应。显著交互作用表明, 增温天数对产量构成因素的影响因品种和果枝而异。在上部果枝中, 铃数在增温水平以及增温水平与品种的交互作用中达到显著水平; 单铃重也在增温水平和品种间达到显著性的差异, 这说明花铃期短期增温对海岛棉生殖生长后期的棉铃发育产生了影响。
高温胁迫能引起棉花生理代谢的絮乱, 包括光合作用下降, 花粉活力降低, 授精不良, 蕾铃脱落率增加, 从而导致产量的下降[25-27]。Oosterhuis等[28]认为, 高温胁迫限制了叶片的光合作用, 减少新陈代谢, 降低了棉花的生长速度。本研究结果表明, 在增温处理期间, 随增温时长的增加, 海岛棉叶片的n呈大幅度下降趋势, 蕾铃大量脱落, 单铃重降低; 品种间, 随增温时间的延长, ‘新海43号’的n下降幅度明显小于‘新海49号’, 这可能是‘新海43号’具有较高耐热性的原因, 对高温如何影响海岛棉的光合生理还需进一步的研究测定。短期胁迫造成的植物生长模式的改变, 决定了营养生长与生殖生长的不平衡性[18,29]。本研究中, 花铃期短期增温延长了棉铃成熟期, 导致单铃重下降, 这与前人[30-31]的研究结果一致。结合棉铃干物质量的累积过程分析认为, 随着增温时间的延长, 棉铃干物质量进入快速积累期滞后, 棉铃发育速度减慢、铃期延长, 尤其是快速积累的终止时间延长。由此可推测, 花铃期短期增温使上部果枝单铃重下降的主要原因是棉铃成熟时间的增加无法弥补棉铃生长速率的降低, 使后期棉铃的生长得不到有效的养分供给。
本研究表明, 在产量构成因素中, 铃数对增温最敏感。因此, 增温主要通过降低单株铃数来降低籽棉产量, 与Bange等[32]的研究结果一致。早期较低位置生殖器官的脱落, 延长了花蕾的产生, 并促使一部分花蕾结铃, 增加晚花率和结铃率, 以补偿早期的产量损失[33]。Sadras[34]和张凯等[35]认为, 增强光合作用能提高作物的光和有效辐射利用率, 这是补偿生长的重要组成部分。本研究中, 增温导致中部果枝棉铃大量脱落, 而上部果枝铃数则显著增加, 且随着增温时间的延长, 上部果枝产量及皮棉贡献率增大, 这表明蕾铃的大量脱落使光合产物更多地向营养器官转移, 叶片衰老缓慢, 进而使后期叶片仍能保持较高的光合速率, 上部果枝产量的增加主要是由于叶片较高的光合能力对上部果枝结铃率有较好的促进作用。Kuai等[36]研究发现, 尽管受短期涝渍胁迫的植株棉铃数量与对照植株相近或者更多, 但顶部果枝棉铃结铃与棉铃开放时间推迟。在许多情况下, 延迟生长的棉铃能将棉铃的生长转移到相对不利的地方, 从而限制纤维素合成的数量与速率, 反过来则会导致纤维品质的降低[37]。因此, 增温造成的蕾铃脱落不仅影响海岛棉的产量, 同时还会影响海岛棉的早熟性与纤维品质。补偿性铃在更高的果枝上是否能维持纤维质量还需要进一步的研究分析。
4 结论
本研究中, 盛花期短期(3~9 d)高温胁迫严重影响了海岛棉的长期生长。盛花期高温胁迫造成大量蕾铃脱落, 降低了叶片n, 导致中部果枝铃数、单铃重、衣分和籽棉产量显著下降。在短期高温胁迫作用下, 生殖生长后期棉铃的发育动态发生了明显的变化。随生育进程的推进, 6~9 d高温胁迫的茎、叶干物质量显著高于对照, 且叶片n的衰减速度减慢, 进而提高了上部果枝的铃数, 增加了上部果枝的籽棉产量。短期高温胁迫降低了棉铃的最终干物质积累量, 减小了最大积累速率, 延长了快速积累期终止时间, 同时降低了上部果枝棉铃的单铃重。‘新海43号’的耐热性高于‘新海49号’, 其主要原因是n、单株结铃数和单铃重下降幅度小且蕾铃脱落率低。这些研究结果有助于为海岛棉选育耐热品种、制定丰产抗逆栽培技术提供理论方法。
[1] LEE J, NADOLNYAK D A, HARTARSKA V M. Impact of climate change on agricultural production in Asian countries: Evidence from panel study[C]//Southern Agricultural Economics Association Annual Meeting. Birmingham, AL: Southern Agricultural Economics Association, 2012: 4–7
[2] 秦大河, STOCKER T. IPCC第五次评估报告第一工作组报告的亮点结论[J]. 气候变化研究进展, 2014, 10(1): 1–6 QIN D H, STOCKER T. Highlights of the IPCC working group Ⅰ fifth assessment report[J]. Progressus Inquisitiones de Mutatione Climatis, 2014, 10(1): 1–6
[3] 任国玉, 初子莹, 周雅清, 等. 中国气温变化研究最新进展[J]. 气候与环境研究, 2005, 10(4): 701–716 REN G Y, CHU Z Y, ZHOU Y Q, et al. Recent progresses in studies of regional temperature changes in China[J]. Climatic and Environmental Research, 2005, 10(4): 701–716
[4] 孔庆平. 我国海岛棉生产概况及比较优势分析[J]. 中国棉花, 2002, 29(12): 19–23 KONG Q P. Production situation and comparative advantage analysis of island cotton in China[J]. China Cotton, 2002, 29(12): 19–23
[5] 曹吉强, 徐红, 李群华. 新疆长绒棉生产调研[J]. 中国棉花, 2011, 38(5): 6–8 CAO J Q, XU H, LI Q H. Investigation on the long-staple cotton production in Xinjiang[J]. China Cotton, 2011, 38(5): 6–8
[6] 田立文, 崔建平, 郭仁松, 等. 新疆长绒棉生产历史回顾与现状分析[A]. 中国农学会棉花分会. 中国农学会棉花分会2016年年会论文汇编[C]. 中国农学会棉花分会:《棉花学报》编辑部, 2016: 7 TIAN L W, CUI J P, GUO R S, et al. Research on history review and actuality analysis of ELS cotton production in Xinjing[A].Cotton Branch of China Agricultural Association. Compilation of papers for the 2016 Annual meeting of Cotton Branch of China Agricultural Association[C].Cotton Branch of China Agricultural Association:Editorial Department of Cotton Journal, 2016: 7
[7] 贺新颖, 周治国, 王友华, 等. 铃期增温对棉花产量、品质的影响及其生理机制[J]. 应用生态学报, 2013, 24(12): 3501–3507 HE X Y, ZHOU Z G, WANG Y H, et al. Effect of increased temperature in boll period on fiber yield and quality of cotton and its physiological mechanism[J]. Chinese Journal of Applied Ecology, 2013, 24(12): 3501–3507
[8] SINGH R P, PRASAD P V V, SUNITA K, et al. Influence of high temperature and breeding for heat tolerance in cotton: A review[J]. Agronomy Journal, 2007, 93: 313–385
[9] 宋桂成, 王苗苗, 曾斌, 等. 高温对棉花生殖过程的影响[J]. 核农学报, 2016, 30(2): 404–411SONG G C, WANG M M, ZENG B, et al. The effects of high-temperature on reproductive process in upland cotton[J]. Journal of Nuclear Agricultural Sciences, 2016, 30(2): 404–411
[10] GUINN G. Abscission of cotton floral buds and bolls as influenced by factors affecting photosynthesis and respiration[J]. Crop Science, 1974, 14(2): 291–293
[11] SNIDER J L, OOSTERHUIS D M, SKULMAN B W, et al. Heat stress-induced limitations to reproductive success in[J]. Physiologia Plantarum, 2009, 137(2): 125–138
[12] KUMAR R, SARAWGI A K, RAMOS C, et al. Partitioning of dry matter during drought stress in rainfed lowland rice[J]. Field Crops Research, 2006, 96(2/3): 455–465
[13] OOSTERHUIS D M. Day or night high temperatures: A major cause of yield variability[J]. Cotton Grower, 2002, 46: 8–9
[14] LOKA D A, OOSTERHUIS D M. Effect of high night temperatures on cotton respiration, ATP levels and carbohydrate content[J]. Environmental and Experimental Botany, 2010, 68(3): 258–263
[15] 马辉, 戴路, 龙朝宇. 高温对棉花生殖器官和纤维发育的影响及应对措施[J]. 中国棉花, 2017, 44(4): 38–39 MA H, DAI L, LONG C Y. Effects of high temperature on cotton reproductive organs and fiber development and countermeasures[J]. China Cotton, 2017, 44(4): 38–39
[16] 孙啸震, 张黎妮, 戴艳娇, 等. 花铃期增温对棉花干物重累积的影响及其生理机制[J]. 作物学报, 2012, 38(4): 683–690 SUN X Z, ZHANG L N, DAI Y J, et al. Effect of increased canopy temperature on cotton plant dry matter accumulation and its physiological mechanism[J]. Acta Agronomica Sinica, 2012, 38(4): 683–690
[17] 余新隆, 易先达. 高温对棉花花药开裂影响的观察[J]. 湖北农业科学, 2004, (2): 39 YU X L, YI X D. Observing the cotton anther slit affected by high temperature[J]. Hubei Agricultural Sciences, 2004, (2): 39
[18] SHI P H, ZHU Y, TIAN Y C, et al. Differential effects of temperature and duration of heat stress during anthesis and grain filling stages in rice[J]. Environmental and Experimental Botany, 2016, 132: 28–41
[19] PETTIGREW W T. Moisture deficit effects on cotton lint yield, yield components, and boll distribution[J]. Agronomy Journal, 2004, 96(2): 377–383
[20] 杨威, 朱建强, 吴启侠, 等. 花铃期短期渍水和高温对棉花叶片光合特性、膜脂过氧化代谢及产量的影响[J]. 棉花学报, 2016, 28(5): 504–512 YANG W, ZHU J Q, WU Q X, et al. The effect of short-term waterlogging and high temperature on photosynthesis, membrane lid peroxidation metabolism, and yield during cotton flowering and boll-forming[J]. Cotton Science, 2016, 28(5): 504–512
[21] 彭世杰, 周仲华, 蒋杰, 等. 花铃期增温对棉花生理指标和产量性状的影响[J]. 作物研究, 2016, 30(2): 123–126 PENG S J, ZHOU Z H, JIANG J, et al. Effects of cotton physiology and yield under warming during the flower and boll stage[J]. Crop Research, 2016, 30(2): 123–126
[22] BROOK K D, HEARN A B, KELLY C F. Response of cotton to damage by insect pests in Australia: Pest management trials[J]. Journal of Economic Entomology, 1992, 85(4): 1356–1367
[23] CONATY W C, TAN D K Y, CONSTABLE G A, et al. Genetic variation for waterlogging tolerance in cotton[J]. Journal of Cotton Science, 2008, 12: 53–61
[24] 薛晓萍, 王建国, 郭文琦, 等. 氮素水平对初花后棉株生物量、氮素累积特征及氮素利用率动态变化的影响[J]. 生态学报, 2006, 26(11): 3631–3640 XUE X P, WANG J G, GUO W Q, et al. Effect of nitrogen applied levels on the dynamics of biomass, nitrogen accumulation and nitrogen fertilization recovery rate of cotton after initial flowering[J]. Acta Ecologica Sinica, 2006, 26(11): 3631–3640
[25] COTTEE N S, BANGE M P, WILSON L W, et al.Developing controlled environment screening for high-temperature tolerance in cotton that accurately reflects performance in the field[J]. Functional Plant Biology, 2012, 39: 670–678
[26] REDDY K R, KAKANL V G, ZHAO D, et al. Interactive effects of ultraviolet-B radiation and temperature on cotton physiology, growth, development and hyperspectral reflectance[J]. Photochemistry and Photobiology, 2004, 79(5): 416–427
[27] SNIDER J L, OOSTERHUIS D M, KAWAKAMI E M, et al. Diurnal pollen tube growth rate is slowed by high temperature in field-grownpistils[J]. Journal of Plant Physiology, 2011, 168(5): 441–448
[28] OOSTERHUIS D M, SNIDER J L. High temperature stress on floral development and yield of cotton[M]. Cordova: The Cotton Foundation, 2011: 1–24
[29] 冯波, 李升东, 孔令安, 等. 灌浆初期高温胁迫对不同耐热性小麦品种形态和产量的影响[J]. 中国生态农业学报, 2019, 27(3): 451–461FENG B, LI S D, KONG L A, et al. Effect of high temperature stress at early grain-filling stage on plant morphology and grain yield of different heat-resistant varieties of wheat[J]. Chinese Journal of Eco-Agriculture, 2019, 27(3): 451–461
[30] NAKANO H, MAKINO A, MAE T. Effects of panicle removal on the photosynthetic characteristics of the flag leaf of rice plants during the ripening stage[J]. Plant and Cell Physiology, 1995, 36(4): 653–659
[31] KATO M, KOBAYASHI K, OGISO E, et al. Photosynthesis and dry-matter production during ripening stage in a female-sterile line of rice[J]. Plant Production Science, 2004, 7(2): 184–188
[32] BANGE M P, MILROY S P, THONGBAI P. Growth and yield of cotton in response to waterlogging[J]. Field Crops Research, 2004, 88(2/3): 129–142
[33] UNGAR E D, WALLACH D, KLETTER E. Cotton response to bud and boll removal[J]. Agronomy Journal, 1987, 79(3): 491–497
[34] SADRAS V O. Compensatory growth in cotton after loss of reproductive organs[J]. Field Crops Research, 1995, 40(1): 1–18
[35] 张凯, 王润元, 王鹤龄, 等. 温度升高和降水减少对半干旱区春小麦生长发育及产量的协同影响[J]. 中国生态农业学报, 2019, 27(3): 413–421 ZHANG K, WANG R Y, WANG H L, et al. Influence of climate warming and rainfall reduction on semi-arid wheat production[J]. Chinese Journal of Eco-Agriculture, 2019, 27(3): 413–421
[36] KUAI J, ZHOU Z G, WANG Y H, et al. The effects of short-term waterlogging on the lint yield and yield components of cotton with respect to boll position[J]. European Journal of Agronomy, 2015, 67: 61–74
[37] ROBERTS E M, RAO N R, HUANG J Y, et al. Effects of cycling temperatures on fiber metabolism in cultured cotton ovules[J]. Plant Physiology, 1992, 100(2): 979–986
Effects of short-term heat stress on island cotton yield formation of different fruiting branches*
CHEN Zhen1, CHEN Ping3, ZHANG Jusong1**, Abudukadier·KUERBAN1, LIN Tao2, GUO Rensong2
(1. Agriculture College, Xinjiang Agricultural University / Research Center of Cotton Engineering, Ministry of Education, Urumqi 830052, China; 2. Institute of Economic Crops, Xinjiang Academy of Agricultural Sciences, Urumqi 830091, China; 3. Awati County Meteorological Bureau, Awati 843200, China)
High temperature is an important environmental factor affecting cotton yield. An experiment was conducted on the effects of short-term heat stress during the full bloom stage on fruit branch yield and yield components of island cotton; it is helpful to provide theoretical methods for breeding heat-resistant and stable-yielding varieties and formulating high-yield and stress-resistant cultivation techniques for island cotton. Using ‘Xinhai 43’ and ‘Xinhai 49’, a warming shed was set up in the field during the full bloom stage, and treatments of four warming levels [lasting for 0 (control), 3 (H3), 6 (H6), and 9 d (H9)] were employed to simulate the effects of short-term high temperature stress. The effects of temperature increase on dry matter accumulation, net photosynthetic rate, abscission rate of squares and bolls, yield, and yield components were analyzed. The results showed that short-term heat stress at the full bloom stage resulted in a significant increase in abscission rate of squares and bolls of the middle fruit branch of ‘Xinhai 43’ and ‘Xinhai 49’, and total dry matter accumulation of cotton plants decreased. At 30–50 days after anthesis, the dry matter of cotton bolls was significantly lower than that of the control. However, at 40–50 days after anthesis, the dry matter of the stems and leaves of two varieties with H6 and H9 treatments were significantly greater than those of the control. As the growth process progressed, compared with the control, the maximum dry matter of ‘Xinhai 43’ and ‘Xinhai 49’ with H3, H6 and H9 treatments decreased by 8.9%, 29.3% and 36.3%, and 11.8%, 28.1%, and 42.6%, respectively; and the daily average accumulation of cotton bolls decreased by 10.9%, 32.8%, and 42.2%, and 12.8%, 30.3%, and 45.9%, respectively. The time needed to reach the terminating date of fleet accumulation period increased by 5, 8, and 14 d, and 1, 5, and 10 d in ‘Xinhai 43’ and ‘Xinhai 49’, respectively. During the warming treatment, thenof the two varieties leaves decreased significantly with longer warming duration. After the end of warming, the senescence rate ofnin each treatment was different. At the later stage of reproductive growth (40 days after anthesis), thenof leaves treated with H3, H6, and H9 was higher than those of the control, and in the order of H9 > H6 > H3 > CK. Short-term high temperature resulted in a significant decrease in boll number per plant, boll weight, and lint percentage. Among them, the coefficient of variation of boll number per plant (15.4% to 18.5%) was the largest, and the coefficient of variation of lint percentage (4.1% to 4.7%) was the smallest. Under the treatments of H3, H6, and H9, the boll number per plant of ‘Xinhai 43’ and ‘Xinhai 49’ decreased by 21.6%, 22.5%, and 28.9% and 16.6%, 26.4%, and 34.7%, respectively. However, the warming significantly increased the number of bolls in the upper fruit branches, and the number of bolls in the upper fruit branches of ‘Xinhai 43’ and ‘Xinhai 49’ increased by 0–24.9% and 2.7%–58.2% compared with the control. This, in turn, increased the yield and yield contribution rate of the upper fruit branches. Among varieties, the decrease inn, boll number, boll weight, and abscission rate of squares and bolls of ‘Xinhai 49’ were greater than those of ‘Xinhai 43’, suggesting that ‘Xinhai 43’ has better heat resistance.
Island cotton; Heat stress; Fruiting branch position; Dry matter accumulation; Leaf net photosynthetic rate; Yield
, E-mail: xjndzjs@163.com
Dec. 20, 2018;
Mar. 26, 2019
S314
2096-6237(2019)09-1375-10
10.13930/j.cnki.cjea.181102
张巨松, 主要研究方向为棉花高产栽培与生理生态。E-mail: xjndzjs@163.com陈振, 主要研究方向为海岛棉丰产抗逆栽培技术及其生理基础。E-mail: 643914079@qq.com
2018-12-20
2019-03-26
* This study was supported by the National Key Research and Development Project of China (2017YFD0101605-05), the National Key Technology R&D Program of China (2014BAD11B02) and the Key Research and Development Project of Xinjiang Uygur Autonomous Region (2016B01001-2).
* 国家重点研发计划项目(2017YFD0101605-05)、国家“十二五”科技支撑计划项目(2014BAD11B02)和新疆维吾尔自治区重点研发项目(2016B01001-2)资助
陈振, 陈平, 张巨松, 阿不都卡地尔·库尔班, 林涛, 郭仁松. 短期高温胁迫对海岛棉不同部位果枝产量形成的影响[J]. 中国生态农业学报(中英文), 2019, 27(9): 1375-1384
CHEN Z, CHEN P, ZHANG J S, Abudukadier KUERBAN, LIN T, GUO R S. Effects of short-term heat stress on island cotton yield formation of different fruiting branches[J]. Chinese Journal of Eco-Agriculture, 2019, 27(9): 1375-1384
