CO2背压啤酒发酵中乙酰辅酶A与酿酒酵母生长和酯类生成的相关性分析
2019-08-30杨东升
杨东升,刘 腊,李 鹏
(海南大学 材料与化工学院,海南 海口 570228)
乙酰辅酶A(acetyl coenzyme A,acetyl-CoA)是酶法合成酯类、脂质、萜类等具有重要工业应用价值的化合物的重要合成前体,参与了微生物中100多条合成与分解代谢途径[1],它既是合成酵母细胞组成物质的关键物质,又是是酯类形成的主要限制因子[2]。乙酰辅酶A是生物合成脂肪酸的基础,也是生物合成酯类的构成物,酯的合成与脂肪酸的合成存在着对酰基辅酶A的竞争机制,因此,有利于脂肪酸的合成,会影响酯的合成[3-4]。酯类是影响啤酒风味最大和最重要的化合物之一,尽管啤酒中的酯类含量不高,但其对啤酒风味的影响非常大。合理控制啤酒中酯类含量,将赋予啤酒协调的风味[5]。
当环境条件有利于酵母的生长繁殖时,例如麦汁中含有足量的可同化氮并在好气条件下,酵母需要合成较多的脂肪酸来合成细胞膜进行繁殖时,酯的合成就会减少。任何影响乙酰辅酶A的生物合成或消耗乙酰辅酶A的反应都会影响酯的生物合成。相反,环境条件不利于酵母的生长繁殖时,如在缺少可同化氮或培养基营养成分不均衡的情况下,酵母较快通过糖酵解形成大量乙酰辅酶A,并以此为底物合成大量酯[6]。特别是啤酒高浓发酵,造成成品啤酒强烈的溶剂味,形成风味缺陷[7]。
CO2是啤酒发酵过程自生产物,在密封的啤酒发酵环境中,其浓度会越来越高直至发酵停止。CO2能影响酵母的代谢途径,改变代谢产物的种类、生成速率和产量[8]。CO2可以直接抑制酵母的生长和代谢,尤其是抑制对酵母代谢起关键作用的脱羧反应,脱羧反应的减少进而影响酯类合成底物乙酰辅酶A的形成[9]。另外,一些酶蛋白上可能存在一个特定的阴离子敏感位点,其会受到HCO-3的抑制[10],使代谢方向发生改变。如在一定压力范围内,随着外加CO2分压的上升,使更多的丙酮酸作为反应底物通过生成乙醛转化成酒精[11-12],从而减少乙酰辅酶A的形成,进而影响酯的生成。研究发现[13],低浓度CO2可以刺激酵母的生长。合理的背压可以控制丙酮酸到乙酰辅酶A的脱羧过程对CO2抑制的敏感度。比如,现代啤酒发酵通过密封罐调整背压确保发酵过程的顺利进行,尤其是大规模的高浓度和超高浓度啤酒发酵技术使发酵环境压力、CO2浓度更高,对酵母生长代谢及酯类代谢的影响更是不能忽视[7]。综上,保持合理的CO2背压非常重要,除了可以控制酵母生长速度,又可以控制乙酰辅酶A的消耗,导致影响酯的形成。然而,现有研究CO2对啤酒生产影响仅局限于单一的因素分析,未能充分揭示在CO2背压下,乙酰辅酶A与酵母生长、酯类形成的相关性。
本研究通过控制比较啤酒背压发酵与常压发酵,通过检测主发酵过程中乙酰辅酶A、酵母生长和酯类形成的变化,分析CO2对啤酒发酵基本参数的影响,揭示乙酰辅酶A与酿酒酵母生长和酯类生成的规律,对指导生产适宜风味的啤酒具有十分重要的作用。
1 材料与方法
1.1 材料与试剂
酿酒酵母(Saccharomyces cerevisiae)NCYC 1108、麦汁(12°Bx):海南大学生物工程综合实验室;柠檬酸合酶(100 U/m L):美国Sigma公司;疏基乙醇、苯甲基磺酰氟(phenylmethylsulfonyl fluoride,PMSF)、甘油、Tris-HCl(pH=7.5)、苹果酸脱氢酶(30.4 kU/m L)、L-苹果酸、氧化型辅酶I(β-nicotinam ide adenine dinucleotide trihydrate,NAD)、聚乙烯吡咯烷酮(polyvinylpyrrolidone,PVP)(均为分析纯):北京索莱宝科技有限公司;乙酸乙酯、乙酸异戊酯、己酸乙酯(均为分析纯):阿拉丁试剂(上海)有限公司。
1.2 仪器与设备
100L啤酒自酿系统:哈尔滨汉德啤酒有限公司;JY92IIN超声波细胞破碎仪:上海沪析实业有限公司;5424高速冷冻离心机:德国Eppendorf公司;6890N气相色谱(gaschromatography,GC)仪:美国Agilent公司;TU-1810紫外可见光分光光度计:北京普析通用仪器有限责任公司;MLS3780高压蒸汽灭菌锅:三洋电机株式会社;ATY224岛津电子天平:日本岛津公司。
1.3 实验方法
1.3.1 啤酒常压发酵和CO2背压发酵工艺流程及操作要点

CO2背压发酵:关闭排空阀,高泡期形成后,使压强自动上升至0.05MPa,保持,如果压力继续升高,则通过排气保持压力,发酵温度10℃。
常压发酵:排空阀保持常开,发酵温度10℃。
1.3.2 分析检测
(1)酵母计数[14]
酵母计数采用血球计数板法。

(2)乙酰辅酶A含量测定[15]
乙酰辅酶A提取试剂:聚乙烯吡咯烷酮(PVP)2.5 g、100mmol/L的巯基乙醇37μL、100mmol/L的苯甲基磺酰氟(PMSF)500μL和甘油5m L溶于50mmol/L,pH值为7.5,100m L的Tris-HCl缓冲溶液后组成,置于试剂瓶中,4℃保存[15]。
乙酰辅酶A检测试剂:苹果酸脱氢酶0.3mg,柠檬酸合酶10 μL,苹果酸30mg和氧化型辅酶I(NAD)7.5mg溶于50mmol/L,pH值为7.5,23m L的Tris-HCl缓冲溶液中[15]。
样品制备:取20 m L发酵液于50 m L无菌离心管中,5 000 r/min离心15min,收集离心沉淀,用生理盐水洗涤酵母菌体两次。弃上清,沉淀按照每400万细菌或培养细胞加入1m L的乙酰辅酶A提取试剂,并且采用功率为20W的超声波,对离心管内的细菌或培养细胞超声3 s,间隔10 s,重复30次,破碎细胞,然后采用13 000×g离心条件,4℃离心10m in,取上清,置冰上,用于测定乙酰辅酶A[15]。
乙酰辅酶A含量测定[15]:分光光度计预热30min,用蒸馏水于波长340 nm处调零;取230μL的乙酰辅酶A检测试剂和25μL样本至微量石英比色皿,混匀;立即记录波长340 nm处20 s时的吸光度值A1和80 s时的吸光度值A2;计算吸光度值ΔA=A2-A1;以吸光度值ΔA(x)为横坐标,以标准品浓度(y,nmol/mg)为纵坐标,绘制乙酰辅酶A标准曲线,采用乙酰辅酶A标准曲线回归方程y=1 640x+0.012计算样品中乙酰辅酶A含量。

(3)总酯含量测定
采用顶空气相色谱法测定乙酸乙酯、乙酸异戊酯、己酸乙酯。气相色谱条件如下:采用毛细管HP-NIONAX柱(30m×0.32mm×0.25μm),顶空采样器HP7694E,注入温度225℃;柱温程序:在40℃条件下保持3min,然后在5℃/min增加到90℃;检测器温度300℃;载气为氮气(N2);载气流量30m L/m in;氢气(H2)流量40m L/m in;空气流量400m L/m in;分流比6∶1;样品瓶平衡温度50℃;平衡时间30m in;循环温度60℃;传输线温度65℃,进样量0.5μL。工作站软件PG2070AA[16-17]。根据保留值进行定性分析,利用外标法(标准曲线法)进行定量计算。

1.3.3 统计方法
所有数据均为3次平行实验结果的平均值。相关性分析采用SAS9.0统计软件。其中P<0.01表示两因素极显著相关;P<0.05表示两因素显著相关[18]。
2 结果与分析
2.1 CO2背压与常压啤酒发酵中乙酰辅酶A、酵母细胞数量及总酯含量及减少率的变化
研究常压和CO2背压条件下啤酒发酵中乙酰辅酶A、酵母细胞数量及总酯含量变化,结果见图1。
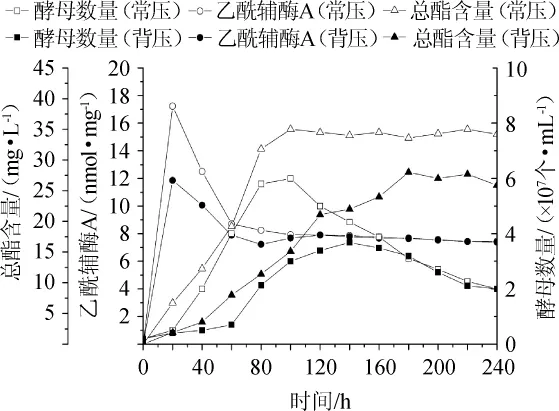
图1 常压与CO2背压啤酒发酵中乙酰辅酶A、酵母细胞数量及总酯含量变化Fig.1 Changes of acetylcoenzyme A,yeast cellnumber and total ester content in beer fermentation under atmospheric pressure and CO2 top pressure conditions
由图1可知,乙酰辅酶A含量在发酵初期(0~20 h)急速增长,并在20 h时达到峰值(常压和CO2背压条件下分别为17 nmol/mg和12 nmol/mg),乙酰辅酶A含量受压力影响明显;发酵时间为20~60 h时乙酰辅酶A含量急剧减少,常压和CO2背压分别减少至8.6 nmol/mg和7.7 nmol/mg;发酵时间为60~100 h,乙酰辅酶A含量缓慢降低,常压和CO2背压分别降低至7.8 nmol/mg和7.5 nmol/mg;发酵时间>100 h之后,常压和CO2背压条件下啤酒发酵中乙酰辅酶A含量均趋于稳定一致。
由图1可知,在常压发酵条件下,酵母细胞数量在0~100 h逐渐增加,并在100 h时达到峰值,为3×107个/m L,发酵时间>100 h之后,酵母细胞数量逐渐下降;在CO2背压发酵条件下,酵母细胞数量在0~140 h逐渐增加,并在140 h时达到峰值,为3.5×107个/m L,发酵时间>140 h之后,酵母细胞数量逐渐下降。两种发酵条件下酵母细胞数量在180 h之后趋于一致。
由图1可知,在常压发酵条件下,总酯的含量在0~100h逐渐增加,并在100 h时达到峰值,为35mg/L,发酵时间>100 h之后,总酯的含量趋于稳定;在CO2背压发酵条件下,总酯含量在0~180 h逐渐增加,并在180 h时达到峰值,为28mg/L,发酵时间>180 h之后,总酯含量趋于稳定。常压发酵条件下总酯含量始终高于CO2背压发酵。
由图1可得初步结论,CO2背压令乙酰辅酶A减少,同时抑制酵母细胞生长及总酯的生成,使酵母细胞生长及总酯含量生成峰值滞后。CO2背压啤酒发酵中乙酰辅酶A、酵母细胞数量及总酯含量减少率结果见图2。

图2 CO2背压引起啤酒发酵中乙酰辅酶A、酵母细胞数量及总酯含量减少率Fig.2 Reduction rate of acetyl coenzyme A,cellnum ber and totalester content in beer fermentation under CO2 top pressure conditions
由图2可知,乙酰辅酶A主要活跃在啤酒发酵0~120h,其中在0~20 h乙酰辅酶减少率上升,并在20 h时乙酰辅酶减少率达到最大,在20~120 h减少率下降,120 h之后减少率稳定。120 h之后对乙酰辅酶A含量影响不大;酵母酵母细胞减少率在0~60 h范围逐渐增大,并在60 h时达到最大,酵母酵母细胞减少率在60~180 h范围急剧减少,酵母酵母细胞减少率在发酵时间>180h之后变化不大;总酯含量减少率在0~20 h范围逐渐增大,并在20 h时达到最大,总酯含量减少率在20~180 h范围逐渐减少,在发酵时间>180 h之后变化不大。CO2背压对酯生成的影响从其他研究中可以得到相似的结果,10 d的啤酒压力发酵抑制了酵母的繁殖和一些风味物质的生成。CO2背压对乙酰辅酶A的主要影响时段为0~120h,120h后未见影响;对酵母数量和总酯含量影响的主要时段为0~180 h,180 h后未见影响。
2.2 常压与CO2背压啤酒发酵乙酰辅酶A与酵母数量的相关性
常压与CO2背压啤酒发酵乙酰辅酶A与酵母数量的相关性见图3。
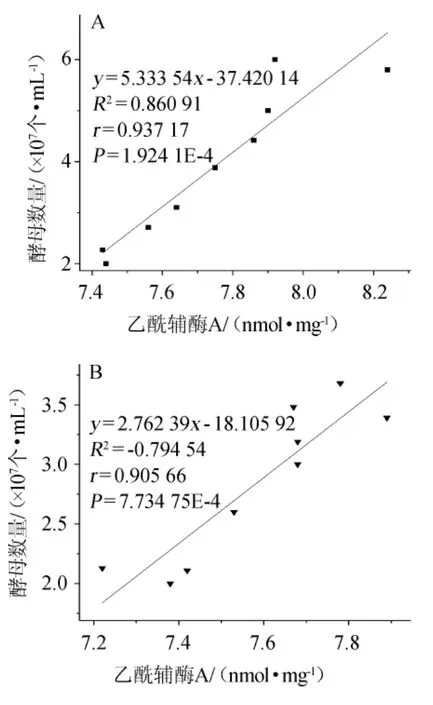
图3 常压(A)和CO2背压(B)啤酒发酵乙酰辅酶A与酵母数量的相关性Fig.3 Correlation between acetyl coenzyme A and yeast cellnumber in beer ferm entation under atm ospheric pressure(A)and CO2 top pressure(B)conditions
由图3A可知,常压条件下啤酒发酵中乙酰辅酶A含量与酵母数量呈极显著正相关(P<0.01),相关系数R=0.93717。由图3B可知,背压条件下啤酒发酵中乙酰辅酶A含量与酵母数量同样呈极显著正相关(P<0.01),相关系数R=0.905 66。
在酵母生长过程中,有很大一部分的碳代谢流通过乙酰辅酶A合成酶途径形成乙酰辅酶A,并参与到其他的细胞器的重要化合物的合成反应中去。因此,乙酰辅酶A的减少将直接影响到酵母数量的减少[5]。
施加CO2背压啤酒发酵后,乙酰辅酶A减少率与酵母数量减少率的相关性见图4。由图4可知,乙酰辅酶A减少率与酵母数量减少率呈显著正相关(P<0.05),相关系数R=0.803 48。说明在CO2背压影响下,酵母数量减少极有可能是乙酰辅酶A减少引起的。酿酒酵母中最丰富的酰基辅酶A是乙酰辅酶A,它可以通过丙酮酸的氧化脱羧或用三磷酸腺苷直接激活乙酸而形成[4]。这是发生在酵母细胞内的反应,在压力的作用下,丙酮酸的氧化脱羧作用受阻[9],减少乙酰辅酶A的生成,同时乙酸受压力影响渗出胞外增加,使得可用于激活的乙酸随之减少[19-20]。酵母生长可利用的乙酰辅酶A减少,细胞数量相应减少。

图4 乙酰辅酶A减少率与酵母数量减少率的相关性Fig.4 Correlation between acetyl coenzyme A reduction rate and yeast cellnumber reduction rate
2.3 常压与CO2背压啤酒发酵乙酰辅酶A与总酯含量的相关性
常压与CO2背压啤酒发酵乙酰辅酶A与总酯含量的相关性见图5。由图5可知,常压条件下啤酒发酵中乙酰辅酶A含量与总酯含量呈显著负相关(P<0.01),相关系数r=-0.83396。CO2背压条件下啤酒发酵中乙酰辅酶A含量与总酯含量呈负相关,相关系数r=-0.750 42。乙酰辅酶A是生物合成脂肪酸的基础,也是生物合成酯类的构成物,酯的合成与脂肪酸的合成存在着对酰基CoA的竞争机制,因此,有利于脂肪酸的合成,会影响酯的合成[3]。换言之,乙酰辅酶A含量的增加有利于酵母生长,会影响酯的合成。
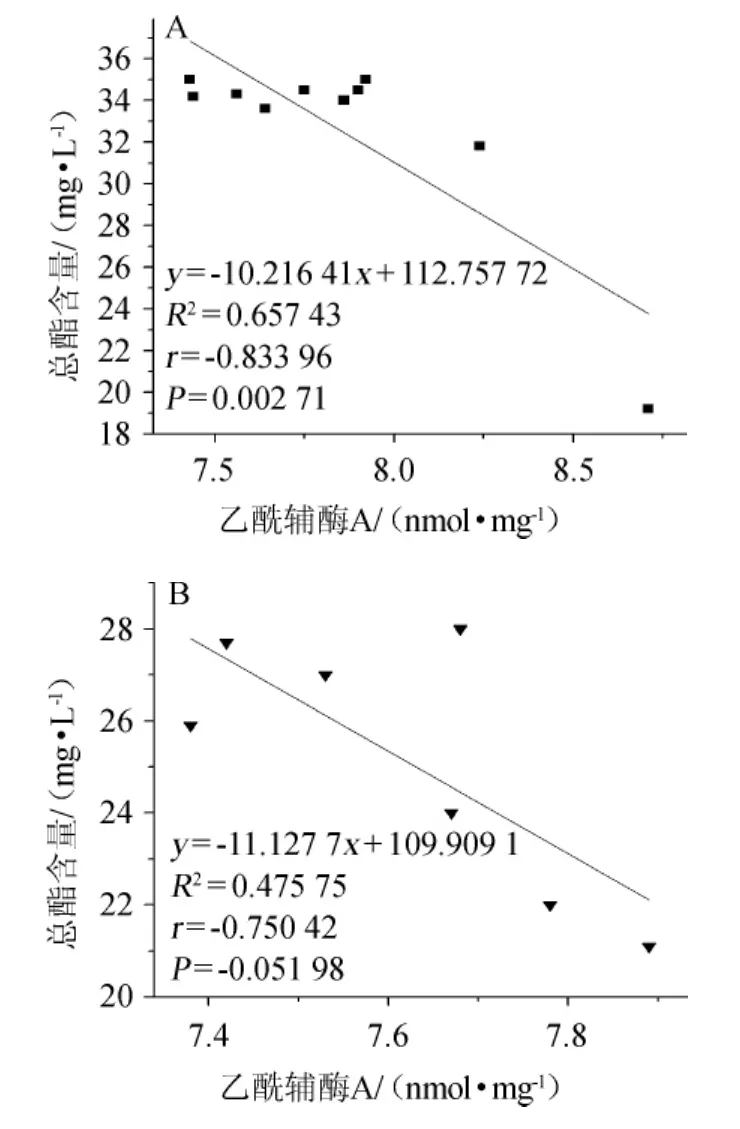
图5 常压(A)和CO2背压(B)啤酒发酵乙酰辅酶A与总酯含量的相关性Fig.5 Correlation between acetyl coenzyme A and totalester content in beer fermentation under atmospheric pressure(A)and CO2 top pressure(B)conditions
乙酰辅酶A的量的分配决定了生化途径的走向和强度,乙酰辅酶A也因此成为一个衡量碳流向的蓄水池,其“水位”的高位决定了酯合成的强度、脂质和氨基酸合成量的大小[4]。结合图1,乙酰辅酶A是酵母细胞生长的先决条件,不管施压与否,乙酰辅酶A的生成总是先于酯的合成。乙酰辅酶A的消耗用于酯的合成,因此两者呈负相关的关系。
施加CO2背压后,乙酰辅酶A减少率与总酯含量减少率的相关性见图6。由图6可知,在CO2背压影响下,乙酰辅酶A减少率与总酯含量减少率呈显著正相关(P<0.01),相关系数r=0.800 29。乙酰辅酶A除了参与细胞内的许多其他反应,包括脂质和氨基酸的生物合成,以及三羧酸循环以外,还参与酯的合成[4]。在乙酰辅酶A的总量控制方面,如果在发酵系统中添加脂质(如长链饱和脂肪酸)就可以减少酵母细胞合成对乙酰辅酶A的需求,增加酯的合成,然而施加CO2背压结果完全相反,乙酰辅酶A的总量因此减少,不单细胞合成受到抑制,酯的合成也受到影响。乙酰辅酶A减少率与总酯含量减少率遵循的这种相关关系,反映了在CO2背压下,乙酰辅酶A对总酯生成的重要性。

图6 乙酰辅酶A减少率与总酯含量减少率的相关性Fig.6 Correlation between acetyl coenzyme A reduction rate and totalester content reduction rate
在正常情况下,酵母需要合成较多的脂肪酸来合成细胞膜进行繁殖时,酯的合成就会减少。任何影响乙酰辅酶A的生物合成或消耗乙酰辅酶A的反应都会影响酯的生物合成。环境条件不利于酵母的生长繁殖时,如在CO2背压的情况下,乙酰辅酶A生成受阻,优先供酵母生长所需,并不会合成大量酯。
3 结论
乙酰辅酶A主要活跃在啤酒发酵0~120 h,乙酰辅酶A含量在发酵初期(0~20h)急速增长,并在20h时达到峰值,常压和CO2背压条件下分别为17 nmol/mg和12 nmol/mg,发酵时间>100 h之后,常压和CO2背压发酵条件下啤酒发酵中乙酰辅酶A含量均趋于稳定一致。常压和CO2背压条件下酵母细胞数量分别在100 h、140 h时达到峰值,两种发酵条件下酵母细胞数量在180 h之后趋于一致。在常压和CO2背压发酵条件下,总酯的含量分别在100 h、180 h时达到峰值,发酵时间>180 h之后,总酯含量趋于稳定。常压发酵条件下总酯含量始终高于CO2背压发酵。常压与CO2背压条件下啤酒发酵中乙酰辅酶A含量与酵母数量均呈极显著正相关(P<0.01)。常压条件下啤酒发酵中乙酰辅酶A含量与总酯含量呈显著负相关(P<0.01),CO2背压条件下啤酒发酵中乙酰辅酶A含量与总酯含量呈负相关。
