不同连作年限烤烟根际土壤真菌群落18S rDNA-PCR-DGGE分析
2019-08-30许自成王发展金伊楠王蒙蒙熊亚南郝浩浩许晓敬
许自成,王发展,金伊楠,王蒙蒙,熊亚南,郝浩浩,许晓敬
(1.河南农业大学烟草学院,河南 郑州 450002;2.河南省烟草公司驻马店市公司,河南 驻马店 463000)
烤烟为忌连作作物,但我国烤烟连作现象普遍存在,导致烟株生长发育不良,抗病抗逆能力降低,烟叶产质量下降,严重影响烟叶生产可持续发展[1-4]。土壤微生物对环境改变和胁迫的反应非常敏感[5],并在土壤内部的变化过程中扮演着重要的角色,因此根际微生物活动是研究烤烟连作障碍的又一重要突破口。近些年来,越来越多的学者认为根际微生态失调可能是连作障碍发生的主要原因并进行大量研究。古战朝等[6]研究发现,根际土壤中细菌的数量随连作年限的增加大致呈“升-降-升-降”的趋势,并跟随烤烟连作年数的延长,根际土壤中细菌所占比例显著降低,真菌比例显著增加,放线菌比例变化则无明显规律。符建国等[7]认为连作造成土壤的理化性质恶化并使植物根系分泌化学物质产生自毒作用,这影响了土壤微生物群落的多样性。罗文富等[8]通过对云南烟区烟草疫霉的研究,认为连作烟田病菌密度高于轮作烟田,可能是随着连作年限的增加,细菌的数量减少,真菌的数量增加。但是,目前有关连作植烟土壤微生物多样性的研究大多集中在连作对土壤微生物数量和细菌群落结构的影响方面[9-12],针对不同连作年限对烤烟根际土壤真菌群落结构的影响的研究较为有限。而且,多数研究主要采用传统微生物平板培养法,利用分子生物学手段较少[13]。平板培养法能较为直观地统计微生物的数量,但只能反映极少数微生物的信息[14]。因此,本研究以河南漯河烟区为例,采用变性梯度凝胶电泳(DGGE)技术并结合真菌的18S rDNA基因测序研究不同连作年限对烤烟根际土壤真菌群落结构和多样性的影响,以期揭示连作植烟土壤微生物群落多样性与结构变化特征,为进一步完善烤烟连作障碍理论提供科学依据。
1 材料与方法
1.1 试验区概况及样品采集
试验于2014年在河南省漯河市郾城区烤烟生产基地(东经114°00′,北纬33°58′)进行。供试烤烟品种为豫烟10号,由漯河市烟草公司提供。该地区植烟土壤为淋溶性黄褐土,土壤基础养分含量为有机质8.12~11.31 g/kg,pH值 7.62~7.83,碱解氮72.6~80.3mg/kg,有效磷15.4~26.1 mg/kg,速效钾113.2~169.5mg/kg。年均气温为14.7℃,年均日照时数2181h,全年无霜期为216~225d,年均降水量为786mm。
试验设3个处理,分别为烤烟轮作(前茬为玉米)、烤烟连作3年和烤烟连作6年。采用大区对比设计进行比较,每个处理面积为2 000 m2,种烟行距、株距分别为110、60 cm。各处理采用“S”型采集法,分别在烤烟团棵、旺长、成熟3个时期,在近株距10cm处,采集0~20cm土壤,重复多次混匀,剔除碎石子及植物残体等,用无菌袋保存于-80℃带回实验室,用于DGGE的测定分析。土壤编号分别为Ⅰ-V1(轮作团棵期)、Ⅰ-V2(连作3年团棵期)、Ⅰ-V3(连作6年团棵期)、Ⅱ-V1(轮作旺长期)、Ⅱ-V2(连作3年旺长期)、Ⅱ-V3(连作6年旺长期)、Ⅲ-V1(轮作成熟期)、Ⅲ-V2(连作3年成熟期)和Ⅲ-V3(连作6年成熟期)。
1.2 真菌DGGE的分析测试
1.2.1 真菌基因组DNA的提取及PCR扩增
本实验土壤总DNA提取采用美国MP Biomedicals生物医学公司生产的FastDNATMSPIN Kit For Soil,提取方法按试剂盒使用说明进行操作。以样品总DNA为模板,采用北京博友顺生物技术有限公司提供的特异性引物GC-Fung(CGC CCG CCG CGC CCC GCG CCC GGC CCG CCG CCC CCG CCC CAT TCC CCG TTA CCC GTT G)/NS1(GTA GTC ATA TGC TFG TCT C)进行扩增[15]。PCR 采用 50μL 反应体系:10× PCR buffer 5μL;dNTP(2.5 mmol/L)3.2μL;rTaq(5U/μL)0.4μL;真菌用 GC-Fung(20 mmol/L)1μL;NS1(20mmol/L)1μL,模板DNA 100 ng;补ddH2O至50μL。PCR扩增程序为:真菌为94℃预变性5min;94℃变性30s、55℃复性30s、72℃延伸30s、30个循环;最终72℃延伸10min。
PCR产物采用OMEGA公司DNA Gel Extraction Kit纯化回收,PCR仪为Biometra公司生产的T-gradient,凝胶成像仪为Bio-Rad公司的Gel-Doc2000凝胶成像系统。
1.2.2 PCR产物的变性梯度凝胶电泳(DGGE)分析
取10μLPCR的产物进行变性梯度凝胶电泳(DGGE)分析[16],真菌采用变性梯度为25%~40%、浓度为8%的聚丙烯酰胺凝胶在1×TAE缓冲液[17]中150V 60℃下电泳8h。变性梯度凝胶电泳(DGGE)完毕后,采用银染法[18]染色。
1.2.3 DGGE图谱中优势条带的回收与测序
用灭菌的手术刀切下待回收DGGE条带,采用OMEGA公司Poly-Gel DNA Extraction Kit回收目的条带,以2μL回收产物为模板,以不带GC夹子的引物进行PCR扩增[19]。
将重新扩增的DNA片段切胶回收、纯化后,连接到pMD18-T载体上,并转化至DH5α感受态细胞中,筛选阳性克隆,菌液由华大基因对插入的真菌18S rDNA片段进行序列测定。
1.2.4 序列分析
将测序所得的真菌18S rDNA优势菌的序列提交至NCBI的GenBank数据库,在GenBank中使用Blast程序进行同源性比较,获得最相似典型菌株的18S rDNA序列。
1.3 统计分析方法
DGGE条带采用Quantity One软件分析;Redundancy analysis(RDA)采用CANOCO 5.0软件进行;图表绘制及数据的整理在Excel 2007软件上进行。
2 结果与分析
2.1 真菌DGGE图谱分析
所得18S rDNA片段通过变性梯度凝胶电泳技术能分离长度相同而碱基组成不同的DNA序列(图1),可以看到明显分离的条带有24条,且多数条带为每个泳道所共有,说明这些处理土壤中大多数真菌类型是共有的。条带位置和数目存在一定差异,说明存在特有的真菌类型。此外,这些公共条带的亮度也有差异,说明土壤真菌在DNA水平上有明显的改变并且真菌种群间的协同演替机制受连作影响。运用Quantity One软件对图谱进行基本的背景排除,经过泳道、条带识别以及配对等步骤得到电泳虚拟图(图2),可以更加直观的看出每个样品的条带差异情况。从条带数量上看,各处理土壤均表现为成熟期>旺长期>团棵期,可见烤烟生育中后期土壤真菌群落结构组成比较复杂。此外,随着生育期的推移,连作的真菌群落变化较轮作明显,且连作6年的变化程度大于连作3年,由此可知不同连作年限对烤烟根际土壤真菌群落有较大影响。
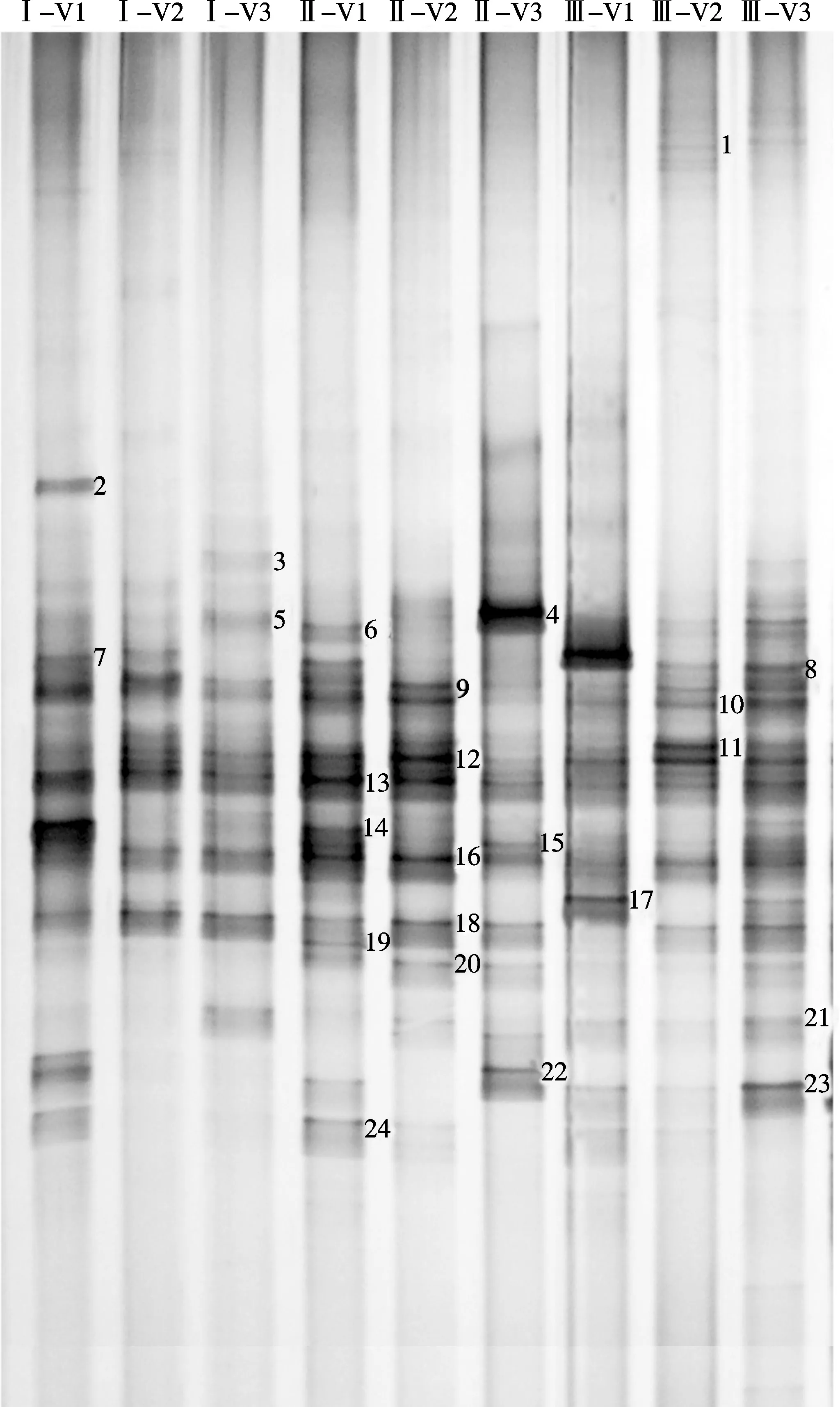
图1 不同连作年限植烟土壤在烤烟大田期真菌DNA图谱

图2 不同连作年限植烟土壤在烤烟大田期真菌DNA电泳虚拟图
2.2 真菌群落相似性指数分析与聚类分析
运用Quantity One 软件对 DGGE 图谱做相似性分析,得到不同处理真菌群落相似性指数(表1)。结果显示,整体上各样品间相似性指数均处于较低水平,表明轮作与连作土壤的真菌群落结构有较大差异,且随着连作年限的延长,相似性是降低的,这说明连作对土壤真菌群落的功能多样性有影响,且随着连作年限的增加影响越大,这与真菌DGGE图谱分析的结果一致。
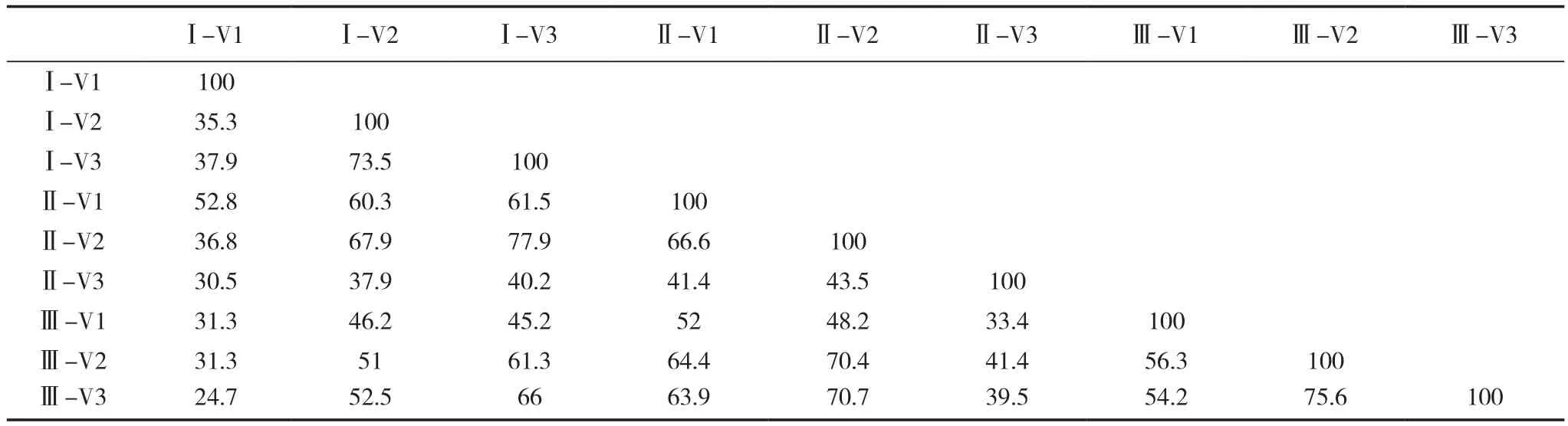
表1 不同连作年限植烟土壤在烤烟大田期真菌群落相似性指数
基于各样本DGGE条带的位置、数量和亮度,采用Quantity One软件对DGGE图谱进行UPGMA聚类分析,见图3。所有样品真菌群落按相似程度可以分为5类,轮作土壤旺长期、连作6年和连作3年的成熟期的真菌群落(Ⅱ-V1、Ⅲ-V3、Ⅲ-V2)能较好聚成一类,连作3年团棵期、连作6年团棵期以及连作3年的旺长期的真菌群落(Ⅰ-V2、Ⅰ-V3、Ⅱ-V2)聚为一类,轮作的团棵期(Ⅰ-V1)、连作6年的旺长期(Ⅱ-V3)、轮作的成熟期(Ⅲ-V1)真菌群落各自为一类,这同样说明连作较大程度地改变了土壤真菌的遗传多样性,且不同时期对真菌群落有较大影响。
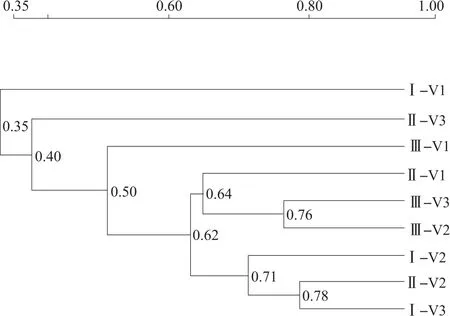
图3 不同连作年限植烟土壤在烤烟大田期真菌群落聚类分析
2.3 真菌群落多样性比较
表2所示,真菌多样性指数在几个样品之间都有所差别。从丰度(S)来看,整体上成熟期>旺长期>团棵期,连作6年>轮作>连作3年。对于均匀度(E),在团棵期,连作6年>连作3年>轮作;在旺长期,轮作>连作3年>连作6年;在成熟期,连作6年>轮作>连作3年,其中最大值出现在连作6年的团棵期,各土壤样品E值有一定差别,也说明了连作改变了真菌群落结构。从香浓指数(H)和辛普森指数来看,整体上各土壤样品在旺长期和成熟期真菌多样性较高,同时期连作6年土壤样品真菌多样性均大于轮作与连作3年土壤,与丰度结果一致。
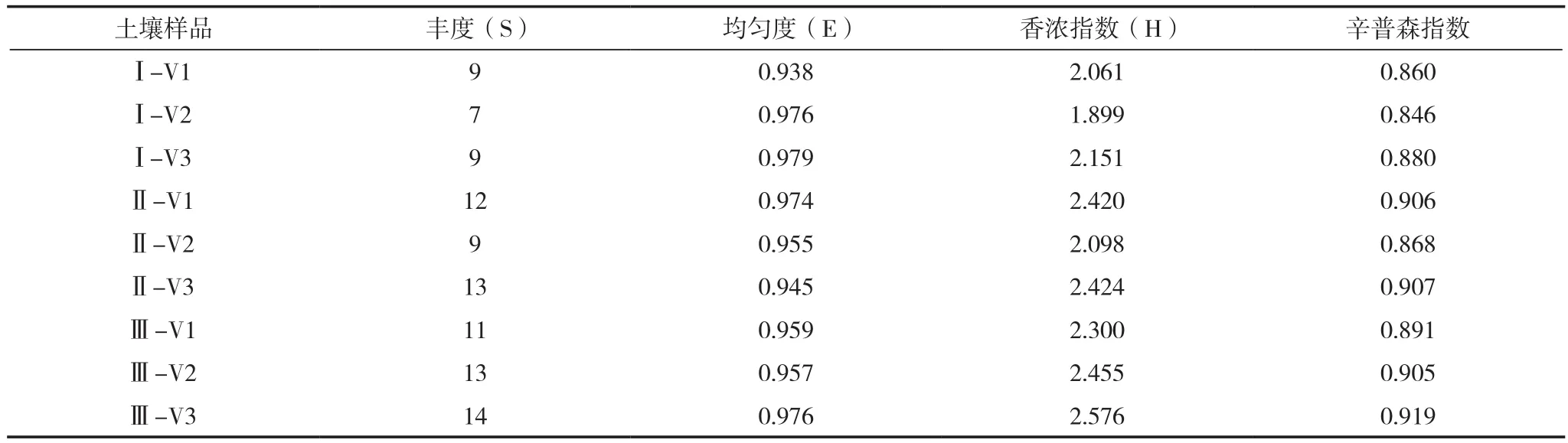
表2 不同连作年限植烟土壤在烤烟大田期真菌多样性指标
2.4 DGGE条带回收序列分析
真菌条带测序后经NCBI比对得到表3。结果表明,从9个样品的DGGE图谱中成功回收的24个条带与数据库中的模式菌株都具有一定的同源性,且相似度在92%~100%,多数与已知菌种的相似度高达97%~100%。一般认为序列同源性小于98%,属于不同种的真菌,同源性小于93%~95%,则属于不同的属[20-21]。因此可以认为真菌菌群优势菌为角担菌科(Ceratobasidiaceae)的立枯菌丝核菌(Rhizoctonia solani)和烟草靶斑病(Thanatephorus cucumeris)、盘菌科(Rhizoctonia)、被孢霉科(Mortierellaceae)、镰刀菌属(Fusarium)、生赤壳科(Bionectriaceae)、漆斑菌属(Myrothecium)、伞菌科(Agaricaceae)、链格孢属(Alternaria)、球盖菇科(Strophariaceae)、炭角菌目(Xylariomycetidae)、链壶菌科(Lagenidiaceae)、腐霉科(Pythiaceae)、花冠菌属(Corollospora)、梳霉科(Kickxellaceae)等。

表3 优势真菌群落的测序克隆序列与其GenBank最相似序列的比对结果
此外,条带9、12、13、16、18存在于烤烟不同生育时期各连作处理土壤中,但条带亮度有不同程度差异,说明其所代表的是烤烟连作土壤中土著的真菌优势种群。而条带11、20和23的真菌种群是连作3年和连作6年土壤中出现过的,条带1的真菌种群仅出现在连作3年的土壤样品中,条带3、4和15的真菌种群仅出现在连作6年的土壤样品中,说明不同连作年限造成一些特异菌群的变化,特别是连作3年和连作6年与轮作相比造成了某些特异性真菌的出现。除此之外,不同生育期植烟土壤真菌结构也有一定差异。条带2只出现于烤烟团棵期的轮作土壤,且亮度清晰可辨,其与立枯丝核菌(Rhizoctonia solani)相似度为99%,可能同属真菌;盘菌科主要分布在连作6年的土壤中,而盘菌科存在于多腐生与腐殖质丰富的土壤中,少数寄生子囊能够引起植物的根腐、茎腐、叶斑等,这也说明了随着连作年限的延长,土壤中病原物质增多。条带7主要存在轮作和连作3年团棵期烟田,与尖孢镰刀菌序列的相似性为100%,说明它在轮作各生育期和连作3年团棵期表现出优势种群;条
带8主要存在于烤烟成熟期轮作、连作土壤中,而其与烟草靶斑病(Thanatephorus cucumeris)之间存在99%的相似度,可能是由于成熟期阴雨天气较多,田间相对湿度较大,有利于靶斑病的发生与蔓延[22]。以上分析可见,烤烟连作后土壤的真菌种群结构发生了变化。
2.5 物种环境因子图
不同连作年限植烟土壤的酶活性和养分含量如表4所示。将不同连作年限植烟土壤的酶活性和养分含量中的土壤蔗糖酶(Invertase)、脲酶(Urease)、碱性磷酸酶(AKP)、蛋白酶(Protease)活性以及土壤碱解氮(AN)、有效磷(AP)、速效钾(AK)含量与真菌条带的相关性做RDA物种因子图(图4),可以看出,纵轴和横轴所能解释的相关变量分别为23.2%和28.1%,分析得知条带1、2、5、8、10、11、15、19均与速效钾含量、蛋白酶活性有较强的正相关关系,而与土壤碱解氮含量、碱性磷酸酶活性、脲酶活性、蔗糖酶活性有较强的负相关关系,条带6、18、24则与之相反,并且条带3、14、17、21、22均与有效磷含量有较强的正相关关系。这说明条带5、8、9、15、17、18、19的菌群活动,对土壤的营养代谢影响较大。根据前述分析可知,条带8、9、15主要存在于连作3年和连作6年的烤烟土壤中,说明烤烟连作后土壤真菌群落结构的变化对土壤的营养代谢有强烈的影响,可能是导致连作后烤烟生长发育受抑的重要原因。
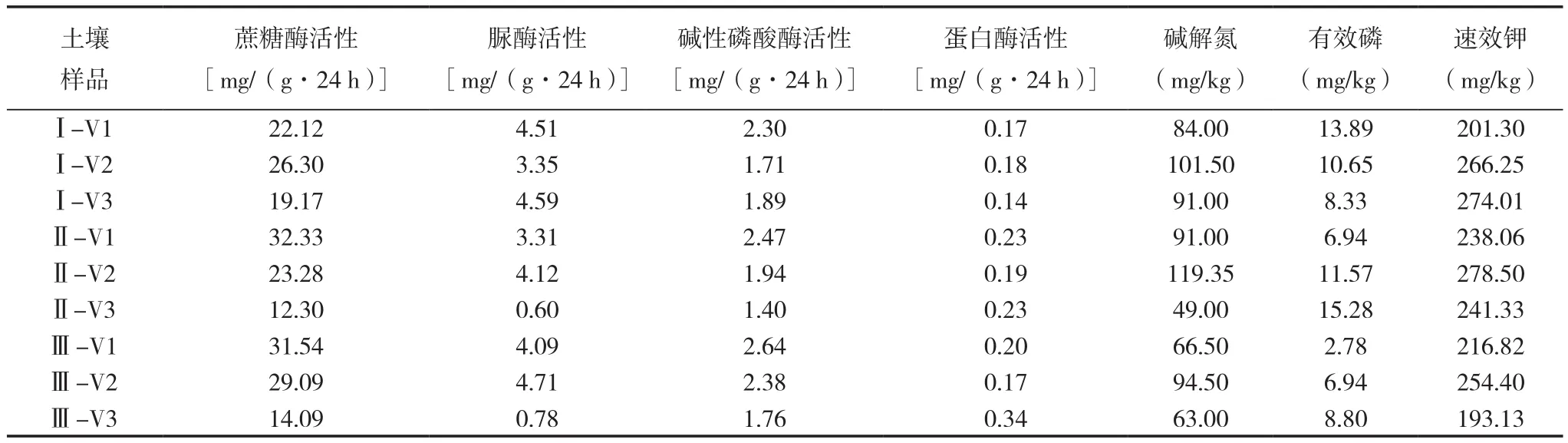
表4 不同连作年限植烟土壤的酶活性和养分含量
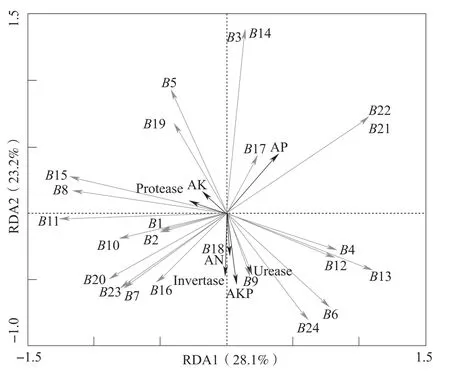
图4 不同连作年限植烟土壤在烤烟大田期优势真菌物种环境因子图
3 结论与讨论
不同连作年限烤烟根际土壤样品在经过 DGGE后,分离出数目不等和迁移率存在明显差异的电泳条带(图1),揭示了不同连作年限烤烟各生育期根际土壤真菌群落的差异性。由电泳图以及多样性指数均能得出,无论是轮作还是连作烤烟生育中后期植烟土壤真菌群落最为丰富,可能是由于成熟期在8~9月份,高温高湿条件下真菌的繁殖与活动增强,并且烟株脱落的烟叶、脚叶落入土壤,为真菌提供充足的营养环境,旺长期则是烟株生长发育最强时期,大量根系分泌物的产生也影响了真菌的繁殖生长[23]。齐虹凌等[24]通过研究轮作与连作对烤烟不同生育期根际土壤细菌群落结构的影响,认为生育期是影响烤烟根际细菌群落结构变化的主要因素,而连作与轮作是次要因素。然而就本试验而言,真菌群落变化对生育期和种植模式的响应程度是否与细菌群落一致尚需进一步地研究。
已有相关研究认为作物连作对土壤真菌群落结构产生了深刻影响[25-26]。本研究的DGGE分析结果表明,不同连作年限烤烟根际土壤样品条带数有明显改变,连作使土壤中真菌种群的个体数明显增多,一些优势种群在不同连作年限的土壤中所处的优势地位发生了很大变化,而且连作6年的变化程度大于连作3年,说明随着连作年限的增加影响越大,这与何川等[9]的研究结果一致。此外,对回收条带序列测序结果表明,不同连作年限造成一些特异菌群的变化,如盘菌科仅出现在连作6年的土壤中,能够引起植物的根腐、茎腐、叶斑等,表明随着连作年限的延长,土壤中病原物质增多。这可能与连作土壤根系分泌物增多、土壤养分失衡等因素有关[27-28]。关于马铃薯连作的研究[19]则表明连作较轮作并未造成马铃薯根际土壤真菌种群数量和多样性的显著变化,这可能与试验区生态环境条件及土壤类型有关,直接表现在作物根际的土著微生物的种类,不同种属或功能型的微生物对连作的敏感性和受性也截然不同;另外还可能与供试作物品种有关,因为其根系分泌物种类和数量均能对根际微生物的活性和组成结构产生影响。有研究表明,连作土壤中某些优势种群富集绝大部分为镰刀菌属等土传病害真菌,主要通过分泌毒素与细胞壁降解酶导致宿主植物发病,并且随着连作年限的增加而逐渐成为优势真菌生理群[29-30],这与本研究结果并不完全一致。从试验结果来看,尖孢镰刀菌属主要存在轮作和连作3年团棵期烟田,而在连作6年土壤中未显示。这可能是随着烟田持续连作,土壤真菌丰富度和多样性不断下降,但连作持续一段时间后土壤微生态系统出现恢复或趋于达到一个新的相对稳定状态,其它真菌群落限制了尖孢镰刀菌属的生存[31]。值得注意的是,土壤漆斑菌属存在于烤烟生长的整个大田期,有研究表明漆斑菌属广泛存在于植物和土壤中,而且大都具有很强的纤维素分解能力,部分菌类还能够产生抗生素[32],土壤漆斑菌属的存在可能对连作土壤微生态的自我修复或化学营养环境的改善有重要影响。总的看来,这些微生物在植烟土壤真菌优势种群平衡中的作用鲜有报道。
微生物群落在土壤有机质分解和营养元素循环中有着至关重要的作用[33]。杨宇虹等[34]指出烟草连作对土壤营养元素循环相关微生物数量有重要影响。土壤微生物群落功能指标与土壤理化性质关系密切,且相互影响。长期连作使土壤的理化性质和生物学性质恶化,导致土壤微生物的多样性和活性下降,从而降低烟草的产质量和加重烟草田间病害[9]。本研究中RDA物种因子图结果显示长期连作导致土壤真菌菌群结构变化,并且烟草靶斑病原与土壤蛋白酶活性有强烈的正相关关系,腐霉科则与土壤速效钾有强烈的正相关性,链壶菌科、生赤壳科的菌群活动与土壤的碱解氮含量、蔗糖酶活性、脲酶活性、碱性磷酸酶活性都有强烈的正相关性,反映了真菌群落与土壤营养元素的关系,但是它们在原位生态环境中的生理生化特性及其所具备的生态功能意义有待进一步的研究。本研究采用的DGGE作为一种分析微生物群落结构组成的分子生物学技术,能够直观地比较和分析微生物群落结构的变化规律,定性地比较土壤微生物群落的差异,尤其是优势微生物类群,但其无法检测基因组中DNA含量少于1.5%的种群,导致不能反映样品中部分低丰度菌种,而高通量测序能提供更多关于低丰度细菌类群的信息[35-36],因此在后续研究中可采用DGGE技术与高通量测序相结合的方法进行,以便能更全面系统地分析不同连作年限土壤样品中真菌群落差异,从而为完善连作土壤中真菌群落结构提供更加全面、真实的信息。
