望江南种子SU5抗菌蛋白的纯化、抑菌机制及稳定性研究
2019-08-23袁素素张玮玮王财成叶秀娟
袁素素,张玮玮,王财成,叶秀娟
(福建农林大学,福建 福州 350002)
苹果黑腐皮壳菌(Valsamali)属于子囊菌亚门黑腐皮壳属,能引起苹果树的树干皮层腐烂,导致苹果树腐烂病[1]。苹果树腐烂病又被称为苹果树癌症,是一种毁灭性病害,严重影响苹果的产量,给农业生产造成巨大的经济损失[2]。该病首次在辽宁省南部区域被发现[3],之后蔓延至我国各个苹果种植区。我国北部地区该病害发生尤为严重,致使苹果树树体布满伤痕,枝干残缺,生长较弱,甚至造成整个种植园的苹果树枯死绝收。但是由于缺乏对苹果树腐烂病流行周期和发病机制的认识,无法对该病害进行有效根本的防治[4]。
望江南,别称野扁豆、金角子或槐豆等,学名为CassiaoccidentalisL.,属1年或多年生豆科决明属植物。在印度,望江南常被用作草药来治疗各种疾病。研究表明,望江南具有抑菌、杀虫、抗癌、抗炎症活性,并能改善胰岛素抵抗状态[5-11]。望江南种子含有的大黄素甲醚吡喃葡糖苷、芦荟大黄素、大黄素甲醚、大黄酚、大黄酸、β-谷甾醇和胡萝卜苷等7种化合物均对HepG2肝癌细胞有一定的抑制活性[10]。分离自望江南的倍半萜2,7-二羟基-4-异丙基-6-甲基萘-1-甲醛和降倍半萜3-异丙基-1,6-二甲氧基-5-甲基萘-7-醇均对癌细胞具有抑制活性[12]。望江南全草甲醇提取物中新蒽醌苷类化合物Emodin-6-O-β-rutinoside和环阿尔屯烷型三萜Cycloccidentaliside C、Cycloccidentalic acid C对人类免疫缺陷病毒(Human immunodeficiency virus,HIV)具有较高的抑制活性[13]。此外,从望江南种子中纯化得到分子质量约为14 ku的杀虫活性蛋白proteinaceous,其也能抑制铜绿假单胞菌和大肠杆菌[7]。然而对望江南种子中抗真菌蛋白及其抗真菌活性的研究却显不足。
本研究选取望江南种子为材料,采用SP-Sepharose阳离子交换层析、Mone S阳离子交换层析和Superdex 75凝胶层析对望江南种子中的抗菌蛋白进行纯化,并通过纸片扩散法、荧光染色法等对此抗菌蛋白的抑菌活性、抑菌机制和稳定性展开研究,以开拓其在苹果黑腐皮壳菌防治中的应用空间,为新型农药的开发提供新的思路和借鉴。
1 材料和方法
1.1 材料
望江南种子:购买于江苏省新泰种业批发有限责任公司。
植物病原真菌:玉米小斑病菌(Helminthosporiummaydis)、尖孢镰刀菌(Fusariumoxysporum)、长丙链格孢菌(Alternarialongipes)、茄病镰刀菌(Fusariumsolanif)、凸脐蠕孢菌(Exserohilummonoceras)、灰葡萄孢菌(Botrytiscinerea)、小白菜炭疽病菌(Colletotrichumhigginsianum)、烟草炭疽病菌(Colletotrichummicotianae)、芭乐炭疽病菌(Colletotrichumgloeosporioides)、稻瘟病菌(Magnaportheoryzae)、落花生球腔菌(Mycosphaerellaarachidicola)和禾谷镰刀菌(Fusariumgraminearum)等12种,均由福建农林大学生物农药与生物化学教育部重点实验室提供。
主要试剂与仪器:SP-Sepharose阳离子交换层析柱、Mone S阳离子交换层析柱和Superdex 75凝胶层析柱(美国GE公司);透析袋(分子截留质量3.5 ku,美国光谱医学公司);4′,6-二脒基-2-苯基吲哚(DAPI)染料(美国Bio-Rad公司);荧光绿染料(Sytox green,美国Cambrex公司);刚果红染料(上海迪柏化学品技术公司);其余化学试剂均为国产分析纯。AKTA purifier蛋白质纯化系统(美国GE公司);LCJ-18S冷冻干燥机(上海松源华兴科技发展有限公司);LDZM立式压力蒸汽灭菌锅(上海申安医疗器械厂);SPH-200F恒温培养摇床(上海世平实验仪器有限公司);Nikon A1激光共聚焦电子显微镜(日本Nikon公司)。
培养基:马铃薯葡萄糖琼脂(PDA)培养基中,马铃薯200 g、葡萄糖20 g、琼脂18 g、蒸馏水1 000 mL;0.7% PDA培养基中,马铃薯200 g、葡萄糖20 g、琼脂7 g、蒸馏水1 000 mL;马铃薯葡萄糖(PD)液体培养基中,马铃薯200 g、葡萄糖20 g、蒸馏水1 000 mL。
1.2 SU5抗菌蛋白纯化
1.2.1 硫酸铵沉淀 将150 g种子清洗干净,加入20 mmol/L NH4OAc缓冲液(pH值4.6) 1 L后,用豆浆机打碎匀浆,4 ℃条件下浸提12 h,然后在10 000 r/min条件下离心30 min,取上清液备用。种子沉淀中再次加入800 mL缓冲液,4 ℃条件下浸提2.5 h,离心收集2次浸提液。在浸提液中加入硫酸铵使饱和度为50%,4 ℃条件下静置4 h后,10 000 r/min离心20 min,取上清液在其中加入硫酸铵使饱和度为100%,4 ℃条件下静置4 h后,10 000 r/min离心20 min,取硫酸铵饱和度为50%~100%时的沉淀蛋白组分。
1.2.2 柱层析纯化 将硫酸铵沉淀蛋白组分溶解在20 mmol/L NH4OAc缓冲液(pH值4.6)中,上样于SP-Sepharose阳离子交换层析柱后,继续用缓冲液洗脱不吸附蛋白,至280 nm吸光值(OD280)为0,得到不吸附蛋白组分,然后换用含有0.2、0.5、1 mol/L NaCl的缓冲液对吸附蛋白依次洗脱。最后以苹果黑腐皮壳菌为指示菌,运用纸片扩散法检测各蛋白质组分的抑菌活性,收集活性蛋白组分,透析并冷冻干燥。
将抑菌活性蛋白组分溶解在10 mmol/L NH4OAc缓冲液(pH值4.6)中,上样于Mone S阳离子交换层析柱后,继续用缓冲液洗脱不吸附蛋白至OD280为0,然后用0~0.3 mol/L NaCl缓冲液对吸附蛋白进行线性盐梯度洗脱。以苹果黑腐皮壳菌为指示菌,检测各洗脱组分抑菌活性,收集活性蛋白组分,透析并冷冻干燥。
将得到的抑菌活性蛋白组分再次溶解在10 mmol/L NH4OAc缓冲液(pH值4.6)中,上样于Superdex 75凝胶层析柱后,用缓冲液洗脱依次得到不同体积的缓冲液洗脱蛋白组分。以苹果黑腐皮壳菌为指示菌,检测各蛋白组分抑菌活性,收集活性蛋白组分,透析并冷冻干燥。
1.2.3 分子质量测定 运用Tricine-SDS-PAGE电泳法,检测蛋白质分子质量。依次配制15%的分离胶、10%的夹层胶和5%的浓缩胶,插入梳子,待凝固后拔出梳子。加入SU5抗菌蛋白样品和蛋白质标准分子质量Marker,在80 V电压条件下进行电泳,待样品进入分离胶后,改用30 V继续电泳至结束。将凝胶取下,置于考马斯亮蓝G-250染色液中染色2 h,然后用脱色液脱色,观察记录电泳结果。
1.3 SU5抗菌蛋白QE质谱鉴定
从Tricine-SDS-PAGE凝胶上切取含目的蛋白的胶条,将样品交由上海中科新生命科技有限公司,通过串联质谱技术,运用Mascot 2.2软件对原始数据进行处理,并检索NCBInr等数据库,鉴定分析蛋白质。
1.4 SU5抗菌蛋白抑菌活性检测
1.4.1 纸片扩散法检测抑菌活性 使用pH值7.4的磷酸盐缓冲液(PBS)配制27.2 μg/mL的SU5抗菌蛋白溶液,备用。将植物病原真菌接种在PDA培养基上,28 ℃恒温培养至菌落直径为3~5 cm时,将直径为6 mm的无菌滤纸圆片放置在距离菌落边缘0.5 cm处,用移液枪吸取15 μL的SU5抗菌蛋白溶液加在滤纸片上,以15 μL的PBS缓冲液为对照,在28 ℃恒温条件下培养至菌落边缘生长至滤纸片,观察记录试验结果。
1.4.2 琼脂稀释法检测抑菌活性 用PBS缓冲液配制2 000 μg/mL的SU5抗菌蛋白溶液,并用0.22 μm滤头过滤除菌,然后吸取300 μL的SU5抗菌蛋白溶液与1 200 μL冷却至50 ℃左右的0.7% PDA培养基混合均匀,倒入培养皿(30 mm×15 mm)中,以300 μL的PBS缓冲液与1 200 μL的0.7% PDA培养基混合为对照,待培养基冷凝后,将直径为5 mm的苹果黑腐皮壳菌菌饼接种在上述培养皿中心位置,最后在28 ℃恒温条件下,培养至对照组菌落长至培养皿边缘处,观察记录试验结果。
1.4.3 孢子萌发抑制活性检测 将苹果黑腐皮壳菌接种于含有PDA培养基的培养皿(90 mm×15 mm)中,28 ℃黑暗条件下培养14 d后,制备1×107cfu/mL孢子溶液,并与0.05%的葡萄糖溶液以体积比1∶1混合。处理组取10 μL孢子溶液和10 μL不同质量浓度的SU5抗菌蛋白溶液混合均匀,对照组取10 μL孢子溶液和10 μL的无菌水混合均匀,之后滴加在凹玻片中央,将凹玻片放入含有2层湿润滤纸的培养皿(90 mm×15 mm)中,28 ℃恒温培养。当对照组的孢子萌发率>90%时,取出凹玻片并置于4 ℃条件下,对每组孢子进行计数,且每组孢子计数总个数不少于100个,每组做3个重复,用SPSS软件对数据进行处理。计算孢子萌发抑制率的公式:孢子萌发率=孢子萌发数/观察孢子数×100%;孢子萌发抑制率=(对照组孢子萌发率-处理组孢子萌发率)/对照组孢子萌发率×100%。
1.5 SU5抗菌蛋白对菌丝尖端几丁质含量、细胞膜透性和DNA降解的影响检测
将苹果黑腐皮壳菌接种于含有PDA培养基的培养皿(90 mm×15 mm)上,待菌落生长至培养皿边缘时,从边缘处切取菌块,放入1.5 mL含有500 μL PD液体培养基的离心管中,处理组中加入500 μL 1772.0 μg/mL的SU5抗菌蛋白溶液,对照组中加入500 μL的PBS缓冲液,在150 r/min、28 ℃恒温条件下培养1 d,然后以1个处理组和1个对照组为一组,分为3组,分别加入刚果红、荧光绿和DAPI染料,并在室温黑暗条件下染色30 min、60 min和30 min,然后用PBS缓冲液洗去多余染液,制作玻片,在荧光显微镜下观察着色结果。
1.6 SU5抗菌蛋白离子稳定性检测
用PBS缓冲液配制100 μg/mL的SU5抗菌蛋白溶液,同时用无菌水分别配制20、100、200、300、400 mmol/L的KCl、MnCl2、FeCl3、CaCl2和MgCl2溶液,然后取等体积的SU5抗菌蛋白溶液与各离子溶液分别混合均匀,以等体积的无菌水和抗菌蛋白溶液混合为阳性对照,以等体积PBS缓冲液和相应400 mmol/L的离子溶液混合为阴性对照,室温放置2 h,最后运用纸片扩散法,以苹果黑腐皮壳菌为指示菌,检测抑菌活性。
2 结果与分析
2.1 SU5抗菌蛋白的纯化
150 g望江南种子经硫酸铵沉淀,得到50%~100%饱和度的沉淀蛋白组分216.36 mg,经检测其对苹果黑腐皮壳菌有抑制活性。将沉淀蛋白组分溶解于20 mmol/L NH4OAc缓冲液(pH值4.6)中,上样于SP-Sepharose阳离子交换层析柱,不吸附蛋白用缓冲液洗脱后,用含有0.2、0.5、1 mol/L NaCl的缓冲液(pH值4.6)进行盐梯度洗脱,分别得到SP1、SP2、SP3和SP4共4个吸附蛋白组分(图1A)。经抑菌活性检测,SP1蛋白组分对苹果黑腐皮壳菌有抑制活性,收集后透析并冷冻干燥,得到SP1蛋白组分39.04 mg。将SP1蛋白组分上样于Mone S阳离子交换层析柱,得到1个未吸附蛋白组分MS0和3个吸附蛋白组分MS1、MS2和MS3(图1B),其中MS3蛋白组分具有抑菌活性,收集透析并冷冻干燥,得到3.79 mg的MS3蛋白组分。将MS3蛋白组分在10 mmol/L NH4OAc缓冲液(pH值4.6)中溶解,上样于Superdex 75凝胶层析柱,经缓冲液洗脱,分别得到SU1、SU2、SU3、SU4、SU5、SU6、SU7和SU8洗脱蛋白组分(图1C)。抑菌检测结果表明,只有SU5蛋白组分对苹果黑腐皮壳菌有抑制活性,为最终的目的蛋白,收集透析后冷冻干燥,得到SU5抗菌蛋白干粉0.44 mg。
采用Tricine-SDS-PAGE电泳检测SU5蛋白组分的分子质量,图1D结果显示,SU5蛋白组分表现为1条带,分子质量约为26 ku,将其命名为SU5抗菌蛋白。

A:SP-Sepharose阳离子交换柱层析;B:Mono S阳离子交换柱层析;C:Superdex 75凝胶柱层析;D:Tricine-SDS-PAGE电泳分析(P:SU5抗菌蛋白;M:蛋白质分子质量Marker)
2.2 SU5抗菌蛋白的QE质谱鉴定
望江南种子SU5抗菌蛋白通过QE质谱鉴定,结果显示,SU5抗菌蛋白为望江南核酮糖-1,5-二磷酸羧基酶/加氧酶(Ribulose-1,5-bisphosphate carboxylase/oxygenase,GenBank登录号:A0A2S0DEZ8)的大亚基蛋白片段,理论相对分子质量为24 667.71。
2.3 SU5抗菌蛋白的抑菌活性
2.3.1 对植物病原真菌的抑制活性 图2显示望江南种子SU5抗菌蛋白对苹果黑腐皮壳菌的抑制效果,与不加入SU5抗菌蛋白的阴性对照相比,加入望江南种子SU5抗菌蛋白后,菌落的直径变小,可知望江南种子SU5抗菌蛋白对苹果黑腐皮壳菌具有抑制活性。进一步通过纸片扩散法检测望江南种子SU5抗菌蛋白对其他植物病原真菌的抑制活性,结果表明,望江南种子SU5抗菌蛋白对玉米小斑病菌、尖孢镰刀菌、长丙链格孢菌、茄病镰刀菌、凸脐蠕孢菌、灰葡萄孢菌、小白菜炭疽病菌、烟草炭疽病菌、芭乐炭疽病菌、稻瘟病菌和落花生球腔菌共11种植物病原真菌具有抑制活性,但是对禾谷镰刀菌无抑制活性。综上,望江南种子SU5抗菌蛋白对植物病原真菌具有较广的抑制活性。

A:培养基含SU5抗菌蛋白的处理组;B:培养基含PBS缓冲液的对照组
2.3.2 对苹果黑腐皮壳菌孢子萌发的抑制活性 通过凹玻片法测定望江南种子SU5抗菌蛋白对苹果黑腐皮壳菌孢子萌发的抑制活性 (图3)。对照组中,孢子几乎全萌发,芽管细长,粗细均匀;与对照组相比,SU5抗菌蛋白质量浓度为1.49 μg/mL时,孢子大多数萌发,芽管伸长,长度较对照组短;SU5抗菌蛋白质量浓度为2.99 μg/mL时,孢子的萌发数锐减,芽管长度变短、畸形;SU5抗菌蛋白质量浓度为5.98 μg/mL时,孢子膨大、畸形,几乎不萌发;抗菌蛋白质量浓度为11.96 μg/mL时,孢子完全不萌发。

SU5抗菌蛋白质量浓度分别为A:11.96 μg/mL;B:5.98 μg/mL;C:2.99 μg/mL;D:1.49 μg/mL;E:0 μg/mL
经统计,SU5抗菌蛋白质量浓度为1.49 μg/mL时,对孢子几乎无抑制作用,伴随着SU5抗菌蛋白质量浓度的增大,孢子萌发抑制率逐渐升高,当质量浓度达到11.96 μg/mL时,SU5抗菌蛋白对苹果黑腐皮壳菌的孢子萌发抑制率为100.00%(表1)。
2.4 SU5抗菌蛋白对菌丝尖端几丁质含量、细胞膜透性和DNA降解的影响
2.4.1 对苹果黑腐皮壳菌菌丝尖端几丁质含量的影响 刚果红是几丁质染料,可与几丁质结合,用来指示望江南种子SU5抗菌蛋白作用下,苹果黑腐皮壳菌菌丝尖端几丁质的积累情况,结果见图4。对照组中,菌丝内未见红色荧光亮点,而在SU5抗菌蛋白处理组中,菌丝尖端可见明显的红色荧光亮点。由此可知,望江南种子SU5抗菌蛋白抑制苹果黑腐皮壳菌时,导致菌丝尖端几丁质的积累,抑制苹果黑腐皮壳菌的生长。

表1 望江南种子SU5抗菌蛋白对苹果黑腐皮壳菌孢子萌发的抑制率Tab.1 The inhibition rate of SU5 antifungal protein from Cassia occidentalis seeds toward spore germination of Valsa mali

P:SU5抗菌蛋白处理组;CK:PBS缓冲液对照组。下同
2.4.2 对苹果黑腐皮壳菌菌丝细胞膜透性的影响 荧光绿可穿过胞膜受损的细胞,使核酸着色,用来检测望江南种子SU5抗菌蛋白作用时,对苹果黑腐皮壳菌菌丝细胞膜透性的影响,结果见图5。对照组中,菌丝内未检测到绿色荧光,而SU5抗菌蛋白处理组的菌丝内,可检测到明显的绿色荧光亮点,因而,望江南种子SU5抗菌蛋白作用于苹果黑腐皮壳菌时,菌丝细胞膜的完整性被破坏,使得细胞膜透性发生改变,从而抑制菌丝生长。
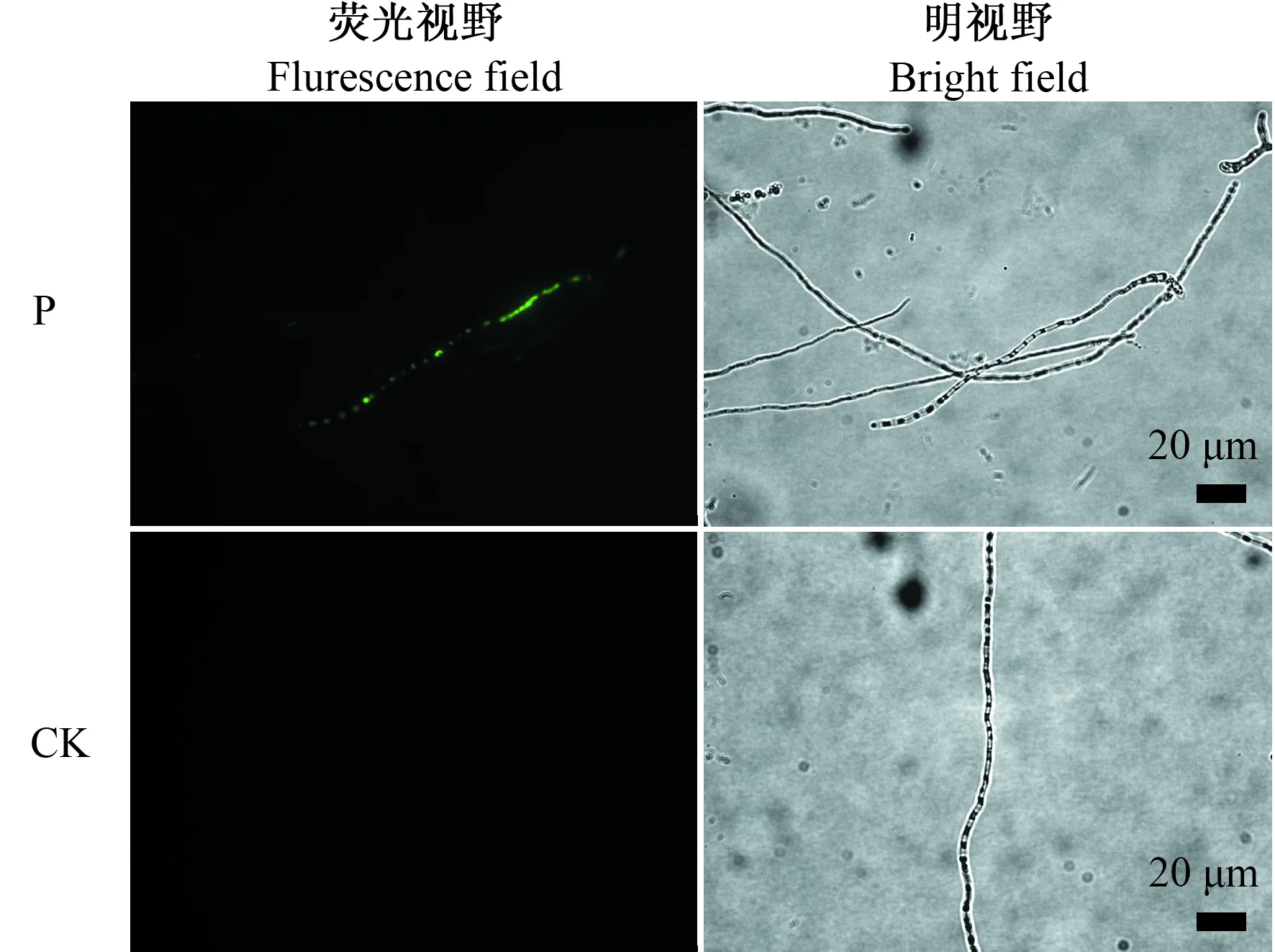
图5 苹果黑腐皮壳菌菌丝荧光绿染色荧光显微镜观察
2.4.3 对苹果黑腐皮壳菌菌丝DNA降解的影响 DAPI是细胞核染料,可与DNA结合,将其作为检测SU5抗菌蛋白对苹果黑腐皮壳菌菌丝DNA降解作用的指示剂,检测结果见图6。对照组中,菌丝内的荧光成规律的圆点状,均匀分布于菌丝内。SU5抗菌蛋白处理组的菌丝内荧光形状不规律,散布于菌丝内。因而,望江南种子SU5抗菌蛋白抑制苹果黑腐皮壳菌时,导致菌丝细胞DNA碎片化、降解,使得菌丝DNA结构被破坏,不规则地分布于菌丝内。
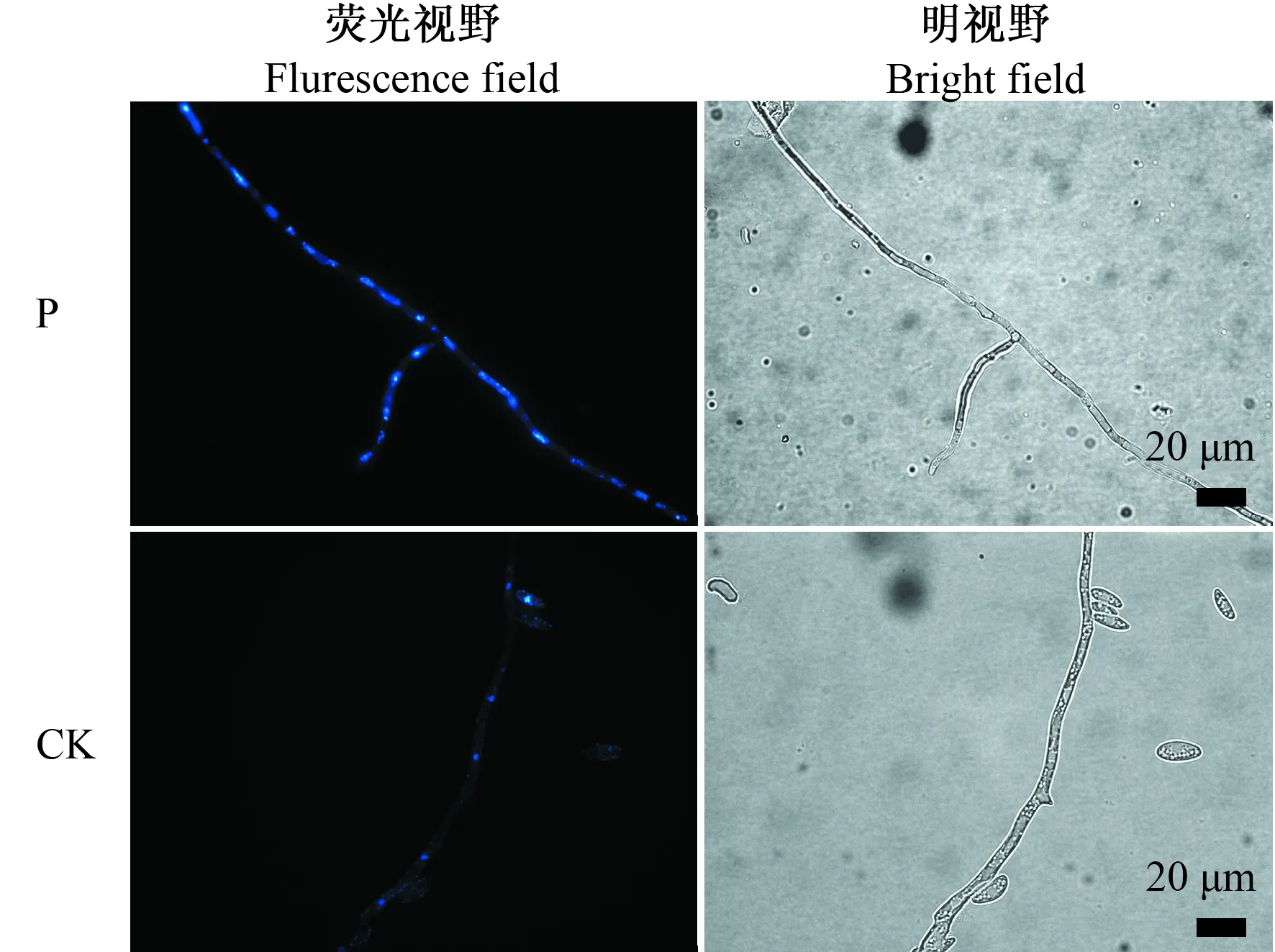
图6 苹果黑腐皮壳菌菌丝DAPI染色荧光显微镜观察Fig.6 Fluorescence microscope images of Valsa mali hyphae stained with DAPI
2.5 SU5抗菌蛋白的离子稳定性
由表2可知,在10~200 mmol/L的K+、Mn2+和Fe3+条件下,SU5抗菌蛋白仍具有抑菌活性;在浓度为10 mmol/L的Ca2+和Mg2+处理后,SU5抗菌蛋白具有抑菌活性,而在Ca2+和Mg2+浓度为50~200 mmol/L时,SU5抗菌蛋白对苹果黑腐皮壳菌的抑制活性消失。综上,K+、Mn2+和Fe3+对望江南种子SU5抗菌蛋白的抑菌活性不产生影响,而Ca2+和Mg2+可抑制SU5抗菌蛋白的抑菌活性。

表2 望江南种子SU5抗菌蛋白在不同离子条件下的稳定性检测结果Tab.2 The stability of SU5 antifungal protein from Cassia occidentalis seeds under different ions conditions
注:+表示对苹果黑腐皮壳菌有抑制活性;-表示对苹果黑腐皮壳菌无抑制活性。
Note:+ represents inhibition activity towardValsamali;- represents no inhibition activity towardValsamali.
3 结论与讨论
望江南种子SU5抗菌蛋白(约26 ku),与桑氏链霉菌KJ07抗菌蛋白的分子质量(28.4 ku)相近[14],而比望江南种子蛋白质提取物(约为14 ku)[8]、芝麻菜种子ZSU2抗菌蛋白(12 ku)[15]和紫菜薹种子抗菌蛋白(13 ku)[16]的分子质量大。
望江南种子SU5抗菌蛋白抑菌活性对K+、Mn2+和Fe3+保持稳定,而被Ca2+和Mg2+所抑制。桑氏链霉菌KJ07抗菌蛋白在Na+、K+、Ca2+、Mg2+、Li+处理后,抑菌活性增加,而经Cu2+、Fe3+、Zn2+处理后,抑菌活性被抑制[14]。紫菜薹种子抗菌蛋白在K+、Zn2+、Mg2+、Cu2+、Pb2+、Al3+、Fe3+和Cr3+金属离子处理后,抑菌活性不受影响,而经Ca2+和Mn2+处理后,抑菌活性被抑制[16]。芝麻菜抗菌蛋白ZSU2在20~150 mmol/L的Ca2+、K+、Mg2+、Mn2+和20~100 mmol/L的Fe3+处理后,抑菌活性不受影响,而在150 mmol/L的Fe3+处理后,其抑菌活性明显减弱[15]。比较发现,望江南种子SU5抗菌蛋白对金属离子的稳定性不如桑氏链霉菌KJ07抗菌蛋白、紫菜薹种子抗菌蛋白和芝麻菜ZSU2抗菌蛋白,其原因可能是望江南种子SU5抗菌蛋白本身是一种酶,而蛋白酶的活性更易受环境影响。
本试验结果表明,SU5抗菌蛋白能抑制12种植物病原真菌,表现出广谱的抑菌活性。VASUDEV等[8]的试验表明,望江南种子蛋白质提取物能抑制烟草赤星病菌、黑斑病菌和尖孢镰刀菌。此外,研究还发现,望江南种子提取物对人体致病细菌和真菌具有很强的抑制活性[17-19]。因而,望江南种子抑菌活性蛋白在实际应用中具有一定的潜力。
望江南种子SU5抗菌蛋白能引起苹果黑腐皮壳菌菌丝尖端几丁质的积累、细胞膜透性发生改变和DNA降解。GHOSH等[20]对大蒜叶凝集素类抗菌蛋白的研究结果表明,抗菌蛋白能够引起立枯丝核菌细胞膜透性改变和DNA降解。紫甘蓝种子抗菌蛋白能引起落花生球腔菌的菌丝尖端出现几丁质积累的现象[21]。
综上,望江南种子SU5抗菌蛋白对植物病原真菌具有一定的抑制活性,但是由于未检测其对具体植物病原真菌的最小抑菌浓度和半抑制浓度,因而不能对其抑菌活性与其他抗菌蛋白进行比较。在后续研究中,需测定望江南种子SU5抗菌蛋白对苹果黑腐皮壳菌的半抑制浓度,并进一步运用分子生物学手段,研究其抑菌机制。
