低温胁迫对山葡萄等6个葡萄品种抗寒性的影响
2019-08-20李桂荣连艳会程珊珊全冉扈惠灵
李桂荣 连艳会 程珊珊 全冉 扈惠灵


摘要:对山葡萄、贝达、北醇、无核白、奥迪亚无核、京可晶这6个葡萄品种的1年生休眠枝条进行不同低温胁迫,测定枝条的相对电导率、超氧化物歧化酶(SOD)活性、过氧化氢酶(CAT)活性、过氧化物酶(POD)活性及可溶性蛋白、可溶性糖、丙二醛、游离脯氨酸含量,并用恢复生长法测定枝条扦插生根率,分析其理化指标变化趋势,应用Logistic方程建立回归曲线模型,拟合其半至死温度(LT50)。结果表明,随胁迫温度的降低,各葡萄枝条SOD、POD活性及游离脯氨酸含量整体呈先升后降趋势,CAT活性呈先升后降再升趋势,相对电导率及可溶性蛋白、可溶性糖、丙二醛含量整体呈上升趋势,扦插枝条生根率呈下降趋势;供试材料的抗寒性强弱依次为山葡萄>贝达>北醇>无核白>奥迪亚无核>京可晶。
关键词:低温胁迫;抗寒性;生理指标;山葡萄;贝达;可溶性糖;可溶性蛋白;丙二醛;脯氨酸
中图分类号: S663.101文献标志码: A
文章编号:1002-1302(2019)08-0130-05
葡萄(Vitis vinifera L.)果实的含糖量相对较高,达到10%~30%,且含多种微量元素,有增进人体健康、治疗神经衰弱和过度疲劳的功效,深受消费者喜爱。目前,我国葡萄栽培多集中在北方地区,但由于北方冬季寒冷,往往导致葡萄冻害频频发生,给许多果农造成巨大的经济损失。因此,有关葡萄的抗寒性成为研究热点,有学者通过测定葡萄相对电导率、防御性酶活性、可溶性糖含量等生理指标来较为系统地研究葡萄抗寒性[1-4]。本试验以无核白、奥迪亚、京可晶这3个无核葡萄品种1年生休眠期枝条为试材,以中国野生山葡萄、砧木品种贝达及山葡萄杂交种北醇为对照,测定低温胁迫下葡萄枝条的电解质渗出率、超氧化物酶(SOD)活性、过氧化物酶(POD)活性、过氧化氢酶(CAT)活性和可溶性蛋白、可溶性糖含量等生理指标及胁迫处理后枝条的扦插生根率,分析其抗寒性强弱,为北方地区葡萄引种、抗寒栽培、种质筛选及抗寒新品种培育提供理论依据。
1材料与方法
1.1试验材料
2016年11月15—30日,从中国农业科学院郑州果树研究所葡萄种质资源圃内剪取山葡萄、贝达、北醇、无核白、奥迪亚、京可晶这6个葡萄品种的中上部1年生休眠枝条,粗 0.5~1.0 cm、长25~50 cm,长势较好,健壮,无病虫害;做好标记,带回沙藏。试验前,将材料取出,清洗数次,置于4 ℃冰箱中保存,备用。
1.2试验设计
试验在河南科技学院园艺园林学院园艺植物遗育种实验室进行,将枝条分别用自来水、去离子水各冲洗3次,滤纸吸干水分;将枝条每个品种平均分成7份,用保鲜膜包好,并用吊牌标记,分别置于4、-5、-10、-15、-20、-25、-30 ℃这7个温度环境下进行胁迫处理,以4 ℃/h的速度降到处理温度,并保持24 h,再以4 ℃/h的速度解冻,每0.5 h回升1个温度梯度;置于4 ℃冰箱中保存,备用,测定其相关指标。重复3次。
1.3测定指标及方法
采用电导率仪测定相对电导率,分别采用氮蓝四唑(NBT)光还原法、愈创木酚法、紫外吸收法测定SOD、POD、CAT活性,分别采用蒽酮比色法、考马斯亮蓝法、茚三酮比色法、硫代巴比妥酸法测定可溶性糖、可溶性蛋白、游离脯氨酸、丙二醛含量[5]。采用曹建东等的方法[6]测定枝条生根率。
1.4数据分析
采用Excel 2010、DPS 15软件对试验数据进行统计分析,采用Origin 8.0软件拟合Logistic方程,并求得半致死温度(LT50)。
2结果与分析
2.1不同低温胁迫对葡萄枝条相对电导率的影响
由图1可见,不同低温胁迫处理对不同葡萄品种的枝条相对电导率有明显影响;随胁迫温度的降低,不同葡萄品种枝条的相对电导率总体呈增加趋势,山葡萄、贝达、北醇枝条的相对电导率变化较为平缓,无核白、奥迪亚、京可晶枝条的相对电导率变化波动较大;奥迪亚、京可晶、无核白葡萄枝条分别在胁迫温度为 -25~-30 ℃、-20~-30 ℃、-20~25 ℃时相对电导率上升较为明显,分别由-25 ℃时的51.4%上升到-30 ℃时的 79.7%、-20 ℃时的55.3%上升到-30 ℃时的76.1%、-20 ℃ 时的50.7%上升到25 ℃时的65.7%;无核白、奥迪亚、京可晶这3个无核葡萄品种的枝条相对电导率在-15 ℃前上升缓慢,随胁迫温度继续降低,枝条相对电导率开始出现跃升,说明这一温度可能已经对细胞膜造成严重伤害,而山葡萄、贝达在温度降到-30 ℃时才有明显上升趋势,说明-15 ℃没有对其造成伤害。因此,山葡萄、贝达的抗寒性相对较强,而无核白、奥迪亚、京可晶这3个无核葡萄品种的抗寒性相对较弱。
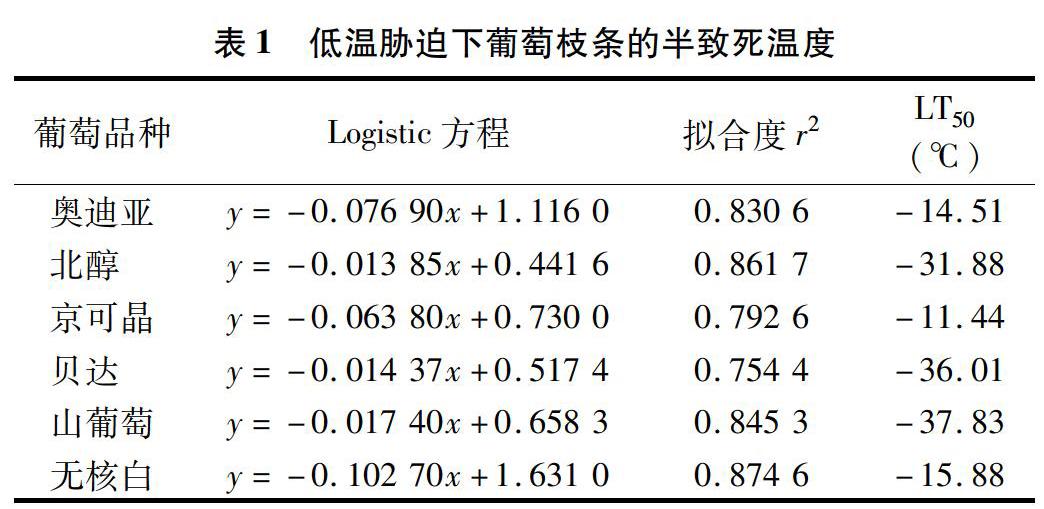
2.2低温胁迫对葡萄枝条半致死温度的影响
对不同温度胁迫下葡萄枝条的相对电导率与相应胁迫温度进行拟合,获得Logistic方程[7-10],并求得半致死温度。半致死温度越高,表明抗寒性越弱,反之,半致死温度越低,表明抗寒性越强。由表1可见,山葡萄、贝达、北醇、奥迪亚、无核白、京可晶的半致死温度(LT50)分别为-37.83、-36.01、-31.88、-14.51、-15.88、-11.44 ℃,表明抗寒性强弱依次为山葡萄>贝达>北醇>无核白>奥迪亚>京可晶。
2.3低温胁迫对不同葡萄枝条酶活性的影响
2.3.1SOD活性由表2可见,随胁迫温度的降低,不同葡萄枝条的SOD活性呈先升后降趋势;山葡萄、贝达、北醇、无核白、京可晶、奥迪亚这6个葡萄枝条的SOD活性在-25 ℃时相对最大,分别为310.05、270.32、264.19、260.74、178.70、208.18 U/g,均极显著高于其他胁迫温度处理(P<0.01),其中,山葡萄的SOD活性在6个葡萄品种中相对最大,贝达次之;[JP3]随胁迫温度的降低,达到-30 ℃时,各葡萄品种枝条SOD活性出现下降,这说明-30 ℃会使葡萄枝條SOD活性可能丧失。
2.3.2POD活性由表2可见,随胁迫温度的降低,不同葡萄枝条的POD活性呈先升后降趋势;山葡萄、贝达、北醇、无核白、京可晶、奥迪亚这6个葡萄枝条的POD活性在-20 ℃时相对最大,分别为16.13、14.56、13.79、13.47、11.52、13.05 U/(g·min),均极显著高于其他胁迫温度处理(P<0.01),其中,山葡萄的POD活性在6个葡萄品种中相对最大,贝达次之;当温度进一步降低,低于-25 ℃时,葡萄枝条POD活性出现持续下降,这说明低于-25 ℃可能会使葡萄枝条POD活性逐渐丧失。
2.3.3CAT活性由表2可见,随胁迫温度的降低,不同葡萄枝条的CAT活性呈先升后降再升趋势;山葡萄、贝达、北醇、无核白、京可晶、奥迪亚这6个葡萄枝条的CAT活性在-20 ℃时相对最大,分别为55.32、47.35、44.39、42.61、32.18、38.74 U/(g·min),均极显著高于其他胁迫温度处理(P<0.01),其中,山葡萄的CAT活性在6个葡萄品种中相对最大,贝达次之。
2.4低温胁迫对葡萄枝条生理指标的影响
2.4.1丙二醛含量在逆境脅迫下,植物器官往往发生膜脂过氧化作用而产生丙二醛,丙二醛对生物膜具有一定的损害作用,其含量越高,对生物膜的伤害程度越大,植物抗逆境胁迫能力相对越弱,反之越强[11]。由表3可见,随胁迫低温的降低,6个葡萄品种枝条的丙二醛含量呈上升趋势;-15 ℃前,随胁迫温度的降低,丙二醛含量上升相对较为缓慢;温度达到-20 ℃时,奥迪亚、京可晶枝条的丙二醛含量出现大幅上升,分别比-15 ℃时增加75.9%、65.8%,山葡萄、贝达上升幅度较小,比-15 ℃时分别增加57.9%、59.1%;-30 ℃低温胁迫时,山葡萄、贝达、北醇、无核白、京可晶、奥迪亚这6个葡萄枝条的丙二醛含量相对最大,分别为0.38、0.39、0.42、0.59、0.68、0.79 μmol/g,多显著高于其他胁迫温度处理(P<0.05),根据丙二醛含量高低可判断各品种枝条耐低温胁迫高低顺序为山葡萄>贝达>北醇>无核白>奥迪亚>京可晶。
2.4.2可溶性糖含量在低温胁迫下,葡萄枝条会对胁迫做出反应,产生大量的糖以提高细胞的渗透浓度、增加原生质浓度,从而降低冰点,进一步提高枝条的抗逆能力。由表3可见,随胁迫低温的降低,6个葡萄品种枝条的可溶性糖含量呈上升趋势;胁迫温度由4 ℃到-15 ℃,各葡萄品种枝条可溶性糖含量缓慢上升,胁迫温度达到-20 ℃时,随胁迫温度的降低,各葡萄品种枝条可溶性糖含量先上升较为迅速;-30 ℃ 低温胁迫时,山葡萄、贝达、北醇、无核白、京可晶、奥迪亚这6个葡萄枝条的可溶性糖含量相对最大,分别为7.32%、6.96%、6.68%、5.44%、5.26%、5.33%,极显著高于其他胁迫温度处理(P<0.01);根据可溶性糖含量高低,可判断各品种枝条耐低温胁迫高低顺序为山葡萄>贝达>北醇>无核 白> 京可晶>奥迪亚。
2.4.3可溶性蛋白含量可溶性蛋白有较强的亲水性,可以使植物细胞的持水力增强,减轻植物体内因原生质结冰而造成的伤害,其含量越高,说明低温胁迫对植物体造成伤害的机率越小,反之越大。由表3可见,随胁迫低温的降低,6个葡萄品种枝条的可溶性蛋白含量整体呈上升趋势;胁迫温度由4 ℃到-25 ℃时,山葡萄、贝达枝条可溶性蛋白含量先上升缓慢,后随胁迫温度降低,可溶性蛋白含量上升较为迅速;胁迫温度由4 ℃到-15 ℃时,奥迪亚、京早晶枝条可溶性蛋白含量先上升缓慢,后随胁迫温度降低,可溶性蛋白含量上升较为迅速;-30 ℃低温胁迫时,山葡萄、贝达、北醇、无核白、京可晶、奥迪亚这6个葡萄枝条的可溶性蛋白含量分别为0.84、0.81、0.72、0.49、0.45、0.46 mg/g,多显著高于其他胁迫温度处理(P<0.05);综合低温胁迫后各葡萄品种枝条可溶[CM(25]性蛋白含量高低,得出各品种枝条耐低温胁迫高低顺序为山葡萄> 贝达>北醇>无核白>奥迪亚>京可晶。
2.4.4游离脯氨酸含量在低温胁迫下,游离脯氨酸含量的增加能维持植物体细胞内环境的稳定,使细胞膜保持结构完整。由表3可见,随胁迫低温的降低,6个葡萄品种枝条的游离脯氨酸含量整体呈先升后降趋势;胁迫温度由4 ℃到 -25 ℃ 时,山葡萄、贝达枝条游离脯氨酸含量先缓慢上升,后随胁迫温度降低,游离脯氨酸含量上升较为迅速;-25 ℃低温胁迫时,山葡萄、贝达葡萄枝条的游离脯氨酸含量相对最大,分别为63.28、59.28 μg/g,-20 ℃低温胁迫时,北醇葡萄枝条的游离脯氨酸含量相对最大,为57.87 μg/g,-15 ℃低温胁迫时,无核白、奥迪亚葡萄枝条的游离脯氨酸含量相对最大,分别为45.83、43.71 μg/g,-10 ℃低温胁迫时,京可晶葡萄枝条的游离脯氨酸含量相对最大,为40.91 μg/g,均相应极显著高于其他胁迫温度处理(P<0.01);综合低温胁迫后各葡萄品种枝条游离脯氨酸含量,得出各品种枝条耐低温胁迫高低顺序为山葡萄>贝达>北醇>无核白>奥迪亚>京可晶。
2.5低温胁迫对葡萄枝条扦插生根率的影响
由图2、 图3可见, 随胁迫低温的降低,各葡萄品种枝条
扦插生根率呈下降趋势;对抗寒性较强的山葡萄、贝达而言,胁迫温度降到-15 ℃时,其生根率仍然保持在100.0%,温度降低到-20 ℃时,其生根率稍有降低,温度降低到-30 ℃时,其生根率分别为45%、38%;对抗寒性较弱的奥迪亚、京可晶而言,胁迫温度降低到-10 ℃时,其生根率分别为33%、29%,已经低于50%,温度下降到-15 ℃时,奥迪亚枝条的生根率仅为5%,而京可晶枝条的生根率为0%。
3结论与讨论
目前,有研究表明,相对电导率、抗氧化酶活性可以较准确地反映植物的抗寒性[12-14]。本试验结果表明,随胁迫温度的降低,不同葡萄品种枝条的电导率总体呈增加趋势,抗寒性较强的山葡萄、贝达、北醇枝条其电导率变化较为平缓,而抗寒性较弱的无核白、奥迪亚、京可晶枝条其电导率变化波动较大;山葡萄、贝达、北醇、奥迪亚、无核白、京可晶这6个葡萄品种枝条的半致死温度分别为-37.83、-36.01、-31.88、-14.51、-15.88、-11.44 ℃,奥迪亚、无核白、京可晶这3个无核品种的半致死温度均在-10~-20 ℃之间,明显低于山葡萄、贝达、北醇这3个对照葡萄品种;葡萄枝条酶活性、生理指标与抗寒性有着密切的关系,随胁迫温度的降低,不同葡萄枝条的超氧化物酶(SOD)、过氧化物酶(POD)活性呈先升后降趋势,过氧化氢酶(CAT)活性呈先升后降再升趋势,这可能是保护酶应对外来胁迫的一种反应,保护酶活性的不断升高,可防御低温对枝条造成的伤害,但达到一定温度时酶活性下降,说明此温度可能导致枝条受到冻害;随胁迫温度的降低,各葡萄品种枝条的丙二醛、可溶性蛋白、可溶性糖含量呈上升趋势,其相应含量高低可在一定程度上可反映各葡萄品种枝条的抗寒性,这与钟海霞等的研究结果[15-17]一致;游离脯氨酸含量整体呈先升后降趋势,这与姜卫兵等的研究结论[18-20]基本吻合。因此,相对电导率、SOD活性、POD活性、CAT活性及可溶性糖、可溶性蛋白、丙二醛、游离脯氨酸含量变化可作为植物抗寒性的鉴定指标,综合试验结果来看,6个葡萄枝条的抗寒性强弱大致为山葡萄>贝达>北醇>无核白>奥迪 亚> 京可晶。
须说明的是,本试验是在人工模拟的低温条件下测定,人工低温处理和室外自然低温会存在一定差异,在今后研究中应结合自然低温条件进行抗寒鉴定,从而使植物抗寒性研究更加准确全面。
参考文献:
[1]贺普超,牛立新. 我国葡萄属野生种抗寒性的研究[J]. 园艺学报,1989(1):81-88.
[2]李淑玲,冯建荣,李亚兰,等. 引入石河子地区的苹果品种抗寒性检测[J]. 果树学报,2012,29(6):1010-1016.
[3]苏李维,李胜,马绍英,等. 葡萄抗寒性综合评价方法的建立[J]. 草业学报,2015,24(3):70-79.
[4]张彪,毛娟,陈佰鸿,等. 葡萄抗寒性鉴定及综合评价方法研究[J]. 甘肃农业大学学报,2014(6):64-69.
[5]邹琦. 植物生理学实验指导[M]. 北京:中国农业出版社,2000:161-173.
[6]曹建东,陈佰鸿,王利军,等. 葡萄抗寒性生理指标筛选及其评价[J]. 西北植物学报,2010,30(11):2232-2239.
[7]李俊才,刘成,王家珍,等. 洋梨枝条的低温半致死温度[J]. 果树学报,2007,24(4):529-532.
[8]曾丽蓉,郑鑫,张婷,等. 电导法协同Logistic方程进行6种苹果砧木抗寒性的比较[J]. 江苏农业科学,2017,45(10):119-121.
[9]邵文鹏,赵兰勇,吴殿鸣,等. 用电导法配合Logistic方程鉴定桉树的抗寒性[J]. 山东林业科技,2009,39(1):28-29.
[10]徐传保,戴庆敏,杨晓琴. 电导法結合Logistic方程确定4种竹子的抗寒性[J]. 河南农业科学,2011,40(11):129-131.
[11]Georgi L,Wang Y,Yvergniaux D,et al. Construction of a BAC library and its application to the identification of simple sequence repeats in peach [Prunus persica (L.)Batsch][J]. Theoretical and Applied Genetics,2002,105(8):1151-1158.
[12]卢精林,李丹,祁晓婷,等. 低温胁迫对葡萄枝条抗寒性的影响[J]. 东北农业大学学报,2015,46(4):36-43.
[13]鲁金星,姜寒玉,李唯. 低温胁迫对砧木及酿酒葡萄枝条抗寒性的影响[J]. 果树学报,2012,29(6):1040-1046.
[14]王敏,陈秋芳,石美娟,等. 低温胁迫对甜樱桃品种枝条SOD、POD活性和脯氨酸含量的影响[J]. 山西果树,2011,8(6):3-5.
[15]钟海霞,艾尔买克·才卡斯木,张付春,等. 7个葡萄砧木根系的抗寒性研究[J]. 新疆农业科学,2016,53(3):429-436.
[16]牛锦凤,李国,王振平. 鲜食葡萄品种抗寒性的比较研究[J]. 陕西农业科学,2005(6):35-36.
[17]何伟,艾军,范书田,等. 葡萄品种及砧木抗寒性评价方法研究[J]. 果树学报,2015,32(6):1135-1142.
[18]姜卫兵,王业遴,马凯. 渗透保护物质在无花果抗寒性发育中的作用[J]. 园艺学报,1992,19(4):371-372.
[19]杨向娜,魏安智,杨途熙,等. 仁用杏3个生理指标与抗寒性的关系研究[J]. 西北林学院学报,2006,21(3):30-33.
[20]尤超,沈虹,郭世荣,等. 早熟油桃品种的抗寒性对比试验[J]. 江苏农业科学,2017,45(14):119-122.
