8种禾本科牧草低温胁迫的生理响应及抗寒性比较
2019-08-20雷雪峰马爱生李海翠王彩艳刘红媛
雷雪峰 马爱生 李海翠 王彩艳 刘红媛


摘要:以来自内蒙古、宁夏、甘肃、新疆、四川、西藏等不同地区的8种禾本科牧草幼苗为材料,分别在15、5、0、-5 ℃ 下对其进行低温胁迫处理,研究其叶片细胞膜透性、丙二醛(MDA)含量、过氧化物酶(POD)活性、游离脯氨酸(Pro)及可溶性蛋白含量等生理生化指标对低温胁迫的响应。结果表明,随着低温胁迫加剧,8种禾本科牧草细胞膜透性、MDA含量总体呈现逐渐上升的趋势,不同物种上升幅度存在差异;而在POD活性、Pro及可溶性蛋白含量方面,不同物种间变化幅度差异较大。通过隶属函数法综合评价得出8种牧草抗寒性表现为细茎冰草>老芒 麦> 沙生冰草>蒙古冰草>多变鹅观草>冰草>黑药鹅观草>大芒鹅观草。
关键词:禾本科牧草;低温胁迫;生理响应;抗寒性
中图分类号: Q945.78文献标志码: A
文章编号:1002-1302(2019)09-0218-05
我国拥有近4亿hm2的天然草地[1-2],但由于全球气候变化及人类对草地不合理的开发利用,已普遍造成不同程度的草地退化,生产力严重下降[3-4]。再加上近年来草地超载过牧严重,草畜矛盾日益加剧,严重地制约着当地畜牧业的快速发展[5]。只靠利用天然草地的养殖方式,已经不适应现代畜牧业生产快速发展的要求[6]。辅助人工草地建设是解决草畜矛盾、改善生态环境的重要手段。在我国北方草原地区,能否正常越冬是制约人工草地建植的关键问题,因此亟需筛选抗寒性较强的牧草品种用于人工草地建植[7]。
禾本科牧草具备产量高、品质好、适口性强、适应性广等特点,在草地畜牧业中发挥重要作用[8]。目前虽然对禾本科牧草抗逆性研究报道较多[9-12],但对其跨区域的种间抗寒性评价研究报道则相对较少。因此,本研究拟对来自内蒙古、宁夏、甘肃、新疆、四川、西藏等不同地区的8种禾本科牧草幼苗进行不同强度的低温胁迫,分析其各相关生理指标的变化情况,通过综合分析,进一步对其进行抗寒性评价,旨在为在我国范围内跨区域挖掘禾本科牧草抗寒材料,以及为寒区人工草地建植适宜品种筛选提供理论依据。
1 材料与方法
1.1 试验材料
供试材料来自于6个地区,包括禾本科3个属8个种,材料具体情况见表1。
1.2 试验方法
1.2.1 试验设计
试验于2016年9月至2017年1月在内蒙古农业大学农学院进行。试验采用盆栽育苗法,苗盆直径为12 cm、高为15 cm,栽培基质为农田土和蛭石,两者体积比为 2 ∶1,苗盆内装入等体积的混合栽培基质,播种量为50粒/盆,盆栽材料全部置于25 ℃(白天)/20 ℃(晚上)的人工气候箱内培养。待苗高达10 cm时进行低温胁迫,胁迫设置4个水平(15、5、0、-5 ℃),并以室温(25 ℃)作对照,每个处理3次重复,各温度胁迫时间为3 d。处理结束后,立刻剪取叶片称质量,并用液氮迅速冷冻,放入-70 ℃低温冰箱中保存备用。
1.2.2 试验测定指标
细胞膜透性测定采用电导仪法;丙二醛(MDA)含量测定采用硫代巴比妥酸显色法;游离脯氨酸[HJ1.4mm](Pro)含量测定采用茚三酮显色法;过氧化物酶(POD)活性测定采用愈创木酚氧化比色法;可溶性蛋白含量测定采用考马斯亮蓝G-250染色法。
1.3 抗寒性综合评价
抗寒性评价采用Fuzzy数学中隶属函数法进行综合分析,其计算公式如下:
与抗寒性呈正相关的参数(POD活性、可溶性蛋白含量
和Pro含量)采用公式(1):
U(Xijk)=(Xijk-Xmin)/(Xmax-Xmin)。(1)
与抗寒性呈负相关的参数(细胞膜透性、MDA含量)采用公式(2):
U(Xijk)=1-(Xijk-Xmin)/(Xmax-Xmin)。(2)
式中:U(Xijk)为第i个禾本科牧草第j个温度阶段第k项指标的隶属度,且U(Xijk)∈[0,1];Xijk表示第i个禾本科牧草第j个温度阶段第k项指标的测定值;Xmax、Xmin分别为所有参试材料中第k项指标的最大值、最小值。运用公式(1)、公式(2)求出不同温度处理下各禾本科牧草各指标参数的隶属函数值,再将其累加起来求其平均值,得出此指標的综合评价值。综合评价值越大,则其抗寒性越强。
1.4 数据分析
用Excel软件进行数据整理和作图,用SPSS 18.0软件进行方差分析、多重比较和抗寒性的隶属函数分析。
2 结果与分析
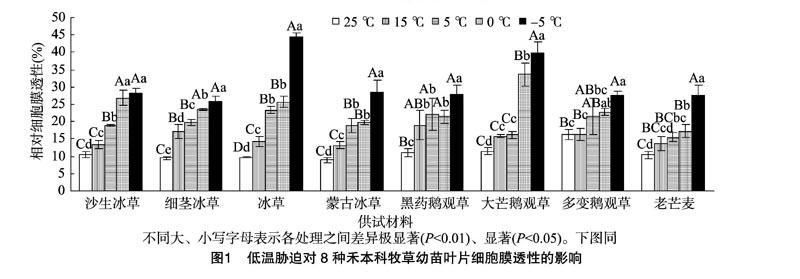
2.1 禾本科牧草幼苗叶片的细胞膜透性变化
如图1所示,随低温胁迫加剧,8种禾本科牧草幼苗叶片细胞膜透性总体呈现逐渐上升的趋势,表明其细胞膜伤害程度逐渐加深。各低温胁迫与对照(25 ℃)相比,8种牧草间幼苗叶片细胞膜透性存在一定差异。在15 ℃条件下,除多变鹅观草及老芒麦外,其他6种牧草叶片细胞膜透性与对照相比,差异均达到显著水平(P<0.05)。在5 ℃条件下,除多变鹅观草外,其他7种牧草幼苗叶片细胞膜透性与对照相比,差异均达到显著或极显著水平。之后,随着胁迫的加剧,各牧草幼苗叶片细胞膜透性与对照相比,差异均达到极显著水平(P<0.01)。其中,在0 ℃条件下,大芒鹅观草叶片细胞膜透性增幅最大,为192.6%,多变鹅观草叶片细胞膜透性增幅最小,为39.0%;在-5 ℃条件下,冰草叶片细胞膜透性增幅最大,为356.6%,多变鹅观草叶片细胞膜透性增幅最小,为68.4%。上述分析表明,低温胁迫对多变鹅观草和老芒麦幼苗叶片细胞膜的伤害程度较小,对冰草和大芒鹅观草幼苗叶片细胞膜的伤害程度较大。
2.2 禾本科牧草幼苗的丙二醛(MDA)含量变化
如图2所示,随低温胁迫加剧,8种禾本科牧草幼苗叶片MDA含量总体呈现逐渐上升的趋势,表明其细胞膜脂过氧化程度逐渐加深。各低温胁迫与对照(25 ℃)相比,8种牧草间幼苗叶片MDA含量存在一定差异。在15 ℃及5 ℃条件下,除沙生冰草、细茎冰草及多变鹅观草外,其他5种牧草叶片MDA含量与对照相比,差异均达到显著或极显著水平。在 0 ℃ 条件下,沙生冰草及多变鹅观草叶片MDA含量与对照相比,差异达到显著(P<0.05)水平,其他6种牧草达到极显著(P<0.01)水平,其中大芒鹅观草和冰草MDA含量的增幅最大,分别为302.0%、200.0%。在-5 ℃条件下,各牧草叶片MDA含量与对照相比均达到极显著(P<0.01)水平,其中,大芒鹅观草和冰草MDA含量的增幅最大,分别为676.4%、490.0%,沙生冰草和细茎冰草MDA含量的增幅最小,分别为216.7%、106.3%。上述分析表明,低温胁迫下沙生冰草和细茎冰草幼苗叶片膜脂过氧化程度较低,大芒鹅观草和冰草幼苗叶片膜脂过氧化程度较高。
2.3 禾本科牧草幼苗的过氧化物酶(POD)活性变化
如图3所示,随着低温胁迫加剧,黑药鹅观草和老芒麦幼苗叶片POD活性呈现逐渐上升的趋势,其他6种牧草幼苗叶片POD活性呈现先上升后下降的趋势,其中细茎冰草、冰草和大芒鹅观草幼苗叶片POD活性峰值出现在胁迫温度为 5 ℃ 时,沙生冰草、蒙古冰草及多变鹅观草幼苗叶片POD活性峰值出现在胁迫温度为0℃时。各低温胁迫处理与对照
(25 ℃)相比,8种牧草间幼苗叶片POD活性存在一定差异。在15 ℃条件下,细茎冰草、黑药鹅观草和多变鹅观草幼苗叶片POD活性与对照相比,差异均达到极显著水平(P<0.01),其他5种牧草与对照相比差异不显著。在5 ℃条件下,8种牧草间幼苗叶片POD活性与对照相比均达到极显著水平(P<0.01)。在0 ℃条件下,细茎冰草和大芒鹅观草幼苗叶片POD活性与对照相比,差异不显著,其他6种牧草与对照相比差异仍到达极显著水平(P<0.01)。在-5 ℃条件下,冰草和大芒鹅观草幼苗叶片POD活性与对照相比,差异不显著,其他6种牧草与对照相比差异仍到达极显著水平(P<0.01)。上述分析表明,低温胁迫下,冰草和大芒鹅观草幼苗叶片POD活性响应较小。
2.4 禾本科牧草幼苗的游离脯氨酸(Pro)含量变化
如图4所示,随低温胁迫加剧,冰草和大芒鹅观草叶片Pro含量呈现先上升后下降的趋势,其他6种禾本科牧草幼苗叶片Pro含量呈现逐渐上升的趋势。各低温胁迫与对照(25 ℃)相比,8种牧草间幼苗叶片Pro含量存在一定差异。在15 ℃条件下,除沙生冰草、细茎冰草、蒙古冰草、大芒鹅观草及多变鹅观草外,其他3种牧草叶片Pro含量与对照相比,差异均达到极显著水平(P<0.01)。在5 ℃条件下,除大芒鹅观草外,其他7种牧草幼苗叶片Pro含量与对照相比,差异均达到极显著(P<0.01)水平。之后,随着胁迫的加剧,各牧草幼苗Pro含量与对照相比,差异均达到极显著水平(P<0.01),其中,在0 ℃条件下,细茎冰草和老芒麦叶片Pro含量增幅最大,分别为173.69%、164.10%,沙生冰草和冰草叶片Pro含量增幅最小,分别为69.50%、65.67%;在-5 ℃条件下,细茎冰草和多变鹅观草叶片Pro含量增幅最大,分别为226.73%、207.50%,冰草和大芒鹅观草叶片Pro含量增幅最小,分别为34.24%、65.36%。上述分析表明,低温胁迫下,沙生冰草、细茎冰草、多变鹅观草和老芒麦幼苗叶片Pro含量响应较大,冰草和大芒鹅观草幼苗叶片Pro含量响应较小。
2.5 禾本科牧草幼苗的可溶性蛋白含量变化
如图5所示,随着低温胁迫加剧,细茎冰草、冰草、蒙古冰草、大芒鹅观草和老芒麦幼苗叶片可溶性蛋白含量总体呈现逐渐上升的趋势,其他3种牧草幼苗叶片可溶性蛋白含量呈现先上升后下降的趋势,其中沙生冰草和多变鹅观草幼苗叶片可溶性蛋白含量峰值分别出现在胁迫温度为5、15 ℃时,黑药鹅观草幼苗叶片可溶性蛋白含量峰值出现在胁迫温度为 0 ℃ 时。各低温胁迫与对照(25 ℃)相比,8种牧草间幼苗叶片可溶性蛋白含量存在一定差异。在15 ℃条件下,除沙生冰草和冰草外,其他6种牧草幼苗叶片可溶性蛋白含量与对照相比,差异均达到极显著水平(P<0.01)。在之后的胁迫温度下,所有材料幼苗叶片可溶性蛋白含量与对照相比,差异均达到极显著水平(P<0.01)。其中,在5、0、-5 ℃条件下,细茎冰草幼苗叶片可溶性蛋白含量增幅均最大,分别为134.3%、153.0%、173.9%,增幅较大的还有蒙古冰草,而冰草(27.9%、50.1%、54.6%)和老芒麦(35.6%、32.4%、61.2%)增幅最小,但其对照基数较大。
2.6 8种禾本科牧草幼苗抗寒性综合评价
植物的抗寒性强弱是多种因子综合作用的结果,因此,为了全面准确地评价供试材料的抗寒性,采用隶属函数法得出各材料各指标的隶属函数值,进一步求得各材料平均隶属度,该值越大,抗寒性越强,反之抗寒性越弱。根据平均隶属度(表2)得出8种禾本科牧草的抗寒性表现为细茎冰草>老芒 麦> 沙生冰草>蒙古冰草>多变鹅观草>冰草>黑药鹅观草>大芒鹅观草。
3 讨论与结论
3.1 禾本科牧草低温胁迫的生理响应
低温胁迫下,植物会发生一系列异常的生理生化反应,其主要表现在细胞膜透性、活性氧含量、抗氧化酶活性及渗透调节物质含量等的改变上,这些变化都是植物受到低温胁迫后的伤害体现或应对反应[13-15]。
低温胁迫对植物的伤害,首先表现为叶片细胞膜系统受损,生物膜透性增加,电解质外渗,电导率增大[16]。同时体内活性氧的平衡被打破,导致膜质过氧化程度加剧,从而膜质过氧化产物丙二醛含量增加。大量研究结果表明,随着低温胁迫强度的增加,早熟禾(Poa annua L.)、黑麦草(Lolium perenne L.)、高羊茅(Festuca elata Keng)[17-18]、老芒麦、偃麦草[Elytrigia repens (L.) Nevski.][19]及其他草坪草[20]等禾本科牧草的相对电导率、细胞伤害率及MDA含量都在增大,只是不同的物种变化程度不同。本研究结果与前人研究结果[17-20]一致,随低温胁迫加剧,供试的8种禾本科牧草幼苗叶片细胞膜透性及MDA含量总体呈现逐渐上升的趋势,表明其细胞膜伤害程度逐渐加深。低温胁迫对多变鹅观草和老芒麦幼苗叶片细胞膜伤害程度较小,对冰草和大芒鹅观草幼苗叶片细胞膜伤害程度较大、膜脂过氧化程度较高。
低温胁迫下,植物体通过调节保护酶活性,清除体内过剩的活性氧自由基,减少有害物质大量积累,进而提高植物抗寒能力[21]。张尚雄等对3个披碱草属(Elymus spp.)牧草的抗寒性研究表明,随着低温胁迫的加剧,保护酶SOD、POD活性随之逐渐增强,不同抗寒性品種间保护酶活性增加幅度不同[22]。但杨顺强等对从美国引进的5种禾本科牧草研究表明,不同温度处理下其根系SOD、CAT、POD的活性变化因种而异,大部分草种均是在冷冻处理后降到最低,返青后才有所回升[23-24]。本研究结果表明,随着低温胁迫加剧,黑药鹅观草和老芒麦幼苗叶片POD活性逐渐上升,其他6种牧草幼苗叶片POD活性呈现先上升后下降的趋势。可见,低温胁迫下,禾本科牧草保护酶活性的变化趋势,与物种的遗传特性、生存环境、低温胁迫的强度以及组织器官等因素有关。
在干旱、低温、高温、盐渍等各种逆境下,植物体均体现出水分胁迫的特征,多数植物可主动积累渗透调节物质来提高细胞液浓度,降低渗透势,提高细胞吸水或保水能力,从而适应逆境胁迫。在抗寒性评价研究中,采用较多的渗透调节物质主要有脯氨酸、可溶性蛋白及可溶性糖,其中可溶性蛋白质含量、脯氨酸含量与低温胁迫关系的研究目前存在2种观点:一种支持低温导致植物细胞中该物质含量变化趋势是逐渐增加的;另一种认为该物质含量在抗冷过程中并不会发生改变,或者与植物抗冷性的增强并不存在因果关系[25]。本研究结果支持前一种观点,即隨着低温胁迫强度加剧,供试材料的渗透调节物质表现为逐渐增加,只是部分材料在低温胁迫后期渗透调节物质增加幅度降低。
3.2 植物抗寒性的综合评价
不同的物种,其遗传特性各异,对环境的适应方式不同,导致其抗寒机制存在差异[26-27]。从报道的文献来看,抗寒性指标主要有形态指标、生理生化指标、分子指标等[28]。但对植物进行抗寒性综合评价时,绝大多数学者还是采用生理生化指标。不同指标对植物抗寒性的贡献率不尽相同,正确评判各指标的贡献率,是植物抗寒性综合评价的关键[29-30]。因此,应采用多个指标综合评价分析植物的抗寒性,才能得出正确的结论。本研究采用了5个生理生化指标,发现随着胁迫强度加剧,均体现出明显的变化规律,研究结果显示与材料抗寒性有较高的相关性,因此在抗寒性综合评价时未进行权重分配。另外,研究发现,在模拟低温胁迫处理时,胁迫方式、梯度、强度对抗寒性的评价结果也有影响,在相关报道中,有直接零下低温胁迫,有零上低温胁迫,有设置回温处理。本研究采用25、15、5、0、-5 ℃的梯度胁迫处理方式,便于观察植物表型变化与低温胁迫强度的关系,可与试验结果相互参比。最终,本研究根据平均隶属度得出8种禾本科牧草的抗寒性表现为细茎冰草>老芒麦>沙生冰草>蒙古冰 草> 多变鹅观草>冰草>黑药鹅观草>大芒鹅观草。
参考文献:
[1]白永飞,王 扬. 长期生态学研究和试验示范为草原生态保护和草牧业可持续发展提供科技支撑[J]. 中国科学院院刊,2017,32(8):910-916.
[2]赵丹丹,马红媛,杨 焜,等. 优质牧草水分利用效率提高技术研究综述[J]. 生态学杂志,2017,36(8):2312-2320.
[3]章力建,王庆锁,侯向阳,等. 中国西部生态农业发展方略[M]. 北京:气象出版社,2004:57-64.
[4]周华坤,赵新全,温 军,等. 黄河源区高寒草原的植被退化与土壤退化特征[J]. 草业学报,2012,21(5):1-11.
[5]侯向阳,尹燕亭,王婷婷. 北方草原牧户心理载畜率与草畜平衡生态管理途径[J]. 生态学报,2015,35(24):8036-8045.
[6]胡自治. 人工草地在我国21世纪草业发展和环境治理中的重要意义[J]. 草原与草坪,2000(1):12-15.
[7]刘 敏,龚吉蕊,张梓瑜,等. 北方干旱区优良牧草抗旱性和抗寒性研究进展[J]. 西北农林科技大学学报(自然科学版),2015,43(3):56-62,76.
[8]王明亚,毛培胜. 中国禾本科牧草种子生产技术研究进展[J]. 种子,2012,31(9):55-60.
[9]马晓林,赵明德,王慧春,等. 高寒牧草在不同温度和盐胁迫作用下的生理生化响应[J]. 生态科学,2016,35(3):22-28.
[10]杨月娟,张 灏,周华坤,等. 盐胁迫对高寒草地牧草老芒麦幼苗生理指标的影响[J]. 西北农业学报,2015,24(7):156-162.
[11]刘锦川,云锦凤. 披碱草属3种牧草幼苗对水分胁迫的响应[J]. 种子,2010,29(9):17-20.
[12]靳军英,张卫华,袁 玲. 三种牧草对干旱胁迫的生理响应及抗旱性评价[J]. 草业学报,2015,24(10):157-165.
[13]Morsy M R,Jouve L,Hausman J,et al. Alteration of oxidative and carbohydrate metabolism under abiotic stress in two rice (Oryza sativa L.) genotypes contrasting in chilling tolerance[J]. Journal of Plant Physiology,2007,164(2):157-167.
[14]刘 璐,毛永成,申亚梅. 3种地被月季对低温胁迫的生理响应[J]. 江苏农业科学,2016,44(2):231-233.
[15]许 娟,郑 虚,闫海锋,等. 不同马铃薯品种苗期叶片对低温胁迫的生理响应[J]. 南方农业学报,2016,47(11):1837-1843. [HJ1.78mm]
[16]姚悦梅,戴忠良,张振超,等. 羽衣甘蓝对低温的生理响应与耐寒性综合评价[J]. 江苏农业学报,2017,33(6):1349-1357.
[17]林立东. 东北高寒地区冷季型草坪草抗寒性研究[J]. 齐齐哈尔大学学报(自然科学版),2008,24(3):83-85.
[18]孙碧徽,李荣全,段小春,等. 多年生黑麦草叶片形态与低温胁迫的生理响应[J]. 草业科学,2016,33(3):450-458.
[19]高丽慧,易 津,李 菲,等. 三种禾本科牧草低温胁迫的生理响应[J]. 中国草地学报,2011,33(3):107-111.
[20]池春玉,丁国华,连永权,等. 低温胁迫对三种冷季型草坪草脯氨酸含量及膜透性的影响[J]. 中国农学通报,2007,23(1):101-104.
[21]赵 慧,赵一博,郭江波,等. 植物耐受低温胁迫研究进展[J]. 种子,2017,36(5):47-50.
[22]张尚雄,尼玛平措,徐雅梅,等. 3个披碱草属牧草对低温胁迫的生理响应及苗期抗寒性评价[J]. 草业科学,2016,33(6):1154-1163.
[23]杨顺强. 不同禾本科牧草在干旱胁迫和低温胁迫下的生理响应[D]. 杨凌:西北农林科技大学,2008:33-34.
[24]李轶冰,杨顺强,任广鑫,等. 低温处理下不同禾本科牧草的生理变化及其抗寒性比较[J]. 生态学报,2009,29(3):1341-1347.
[25]程军勇,郑京津,窦坦祥,等. 植物抗寒生理特性综述[J]. 湖北林业科技,2017,46(5):16-20.
[26]李文明,辛建攀,魏驰宇,等. 植物抗寒性研究进展[J]. 江苏农业科学,2017,45(12):6-11.
[27]钱 稷,张海旺,邸 葆,等. 基于电阻抗图谱法测定桃树抗寒性[J]. 江苏农业科学,2017,45(11):102-104.
[28]李文明,辛建攀,魏驰宇,等. 植物抗寒性研究进展[J]. 江苏农业科学,2017,45(12):6-11.
[29]曹哲群,肖芙荣,陈疏影,等. 7个蔗茅野生种及其后代材料苗期耐寒性鉴定[J]. 作物杂志,2017(5):43-48.
[30]徐呈祥. 植物抗寒性鉴定与测试方法研究进展[J]. 广东农业科学,2014,41(16):50-54.
