单孢继代培养对茄链格孢生长和产孢的影响
2019-08-20刘华娇谷青赵冬梅张岱潘阳杨志辉朱杰华
刘华娇 谷青 赵冬梅 张岱 潘阳 杨志辉 朱杰华
摘要:继代培养对茄链格孢(Alternaria solani)的形态、菌落颜色、生长和产孢等生物学性状均会产生影响。经过连续单孢分离和培养,获得茄链格孢的2次继代培养菌株。随着继代培养代数的增加,角变现象呈上升趋势,F1、F2代菌株出现角变现象的概率分别为23.33%、52.50%。通过十字交叉法和系列稀释法分别测定继代培养菌株的生长情况和产孢量。结果表明,无性F1、F2代与亲代相比,其生长量和产孢量均呈下降趋势,且F2代下降百分率更高。与亲代菌株相比,F1代的生长量下降1.25%~7.61%,F2代生长量下降10.07%~22.29%;F1代菌株单位面积产孢量降低 12.80%~84.60%,F2代菌株单位面积产孢量下降80.11%~96.50%,且产孢量与菌落颜色深浅密切相关,菌落颜色较深的菌株分生孢子单位面积的产孢量要高于菌落颜色浅的菌株。表明茄链格孢无性后代活力不断降低。
关键词:马铃薯早疫病;继代培养;生长量;产孢量;角变
中图分类号: S435.32文献标志码: A
文章编号:1002-1302(2019)09-0160-03
由茄链格孢(Alternaria solani)引起的马铃薯早疫病是影响我国马铃薯生产的重要真菌病害之一。严重时,该病对马铃薯主产区造成的损失可达70%~80%[1]。马铃薯早疫病是一种气传多循环流行性病害,病菌对环境的适应能力强。自茄链格孢被报道以来,前人的研究多集中在其生物性状、侵染方式、药效防治等方面,关于实验室条件下单孢继代培养对茄链格孢生长情况及适应能力的影响国内外未见报道。
研究表明,继代培养可引起虫草、白僵菌等微生物的变异和生物学特性的改变。刘荻等发现,北虫草的组织分离株在继代培养中会发生明显的角变[2]。刘海英等发现,继代培养会造成玉米大斑病菌的菌丝生长速度变慢,产孢量降低[3]。张慧等研究发现,白僵菌产孢量随继代培养代数呈指数下降趋势[4]。许志斌对绿僵菌进行研究发现,随着继代培养代数的增多,菌株的产孢量逐渐降低[5]。孙召朋通过继代培养降低了白僵菌对玉米螟的毒力[6]。唐晓庆等研究发现,继代培养能逐渐增强球孢白僵菌的抗旱能力[7]。
Meng等分析发现,茄链格孢病菌的遗传变异很高,推测其可能存在隐藏的有性生殖[8]。除有性生殖外,还有一种可能是茄链格孢存在准性生殖现象,使病菌的遗传变异增高。单孢分离技术在病原真菌遗传特性、致病力分化等研究中有重要作用。单孢分离可以获得纯菌种,有利于更好地研究菌株的遗传变异。本试验通过对茄链格孢的单孢分离及其后代的继代培养,比较亲代与后代之间生长和产孢的差异,分析继代培养对茄链格孢的影响,旨在为说明茄链格孢较高的遗传变异及是否存在准性生殖现象提供理论依据。
1 材料与方法
本试验于2016年8月至2017年1月完成于河北农业大学马铃薯病害研究中心。
1.1 供试菌株
在笔者所在实验室的菌库中选取6株茄链格孢菌株,分别是采集自河北省的HB-1、HB-6,黑龙江省的HLJ-1、HLJ-9,内蒙古自治区的NMG-1和陕西省的SX-1。
1.2 单孢分离和继代培养
对6株野生型茄链格孢进行产孢培养并进行单孢分离。产孢培养参照刘丽丽的培养方法[9]进行诱导产孢;采用稀释法进行单孢分离,向每皿经诱导产孢的茄链格孢中加入5 mL无菌水进行冲洗,制备成孢子悬浮液,在一次性皿中倒入较薄的一层马铃薯葡萄糖琼脂(PDA)培养基,取1 mL稀释过的孢子悬浮液涂于皿中,静待一段时间,待培养基表面无流动液体时,在显微镜下将周围无菌丝的单个分生孢子连带培养基一起切割转移到新的培养基上培养,获得下一代菌株。
1.3 茄链格孢继代培养菌株生长量的测定
每个菌株分离10株单孢,培养7 d后采用十字交叉法测量菌落直径,计算其平均值。
1.4 茄链格孢继代培养菌株单位面积产孢量的测定
菌株进行產孢培养3 d后,用5 mL无菌水在菌落上反复冲洗,使孢子脱落至无菌水中。将孢子悬浮液收集至10 mL离心管中,统计10倍物镜视野下分生孢子的总数量。每次观察并记录5个视野下的孢子总数,计算其平均值。每个菌株至少检测3皿长势一致的菌落,取其平均值。
单位面积产孢量=每皿的产孢量/每皿的菌落面积。
1.5 数据统计及分析
利用SPSS 19软件对茄链格孢在继代培养中的菌株菌落直径和单位面积产孢量进行方差分析。
2 结果与分析
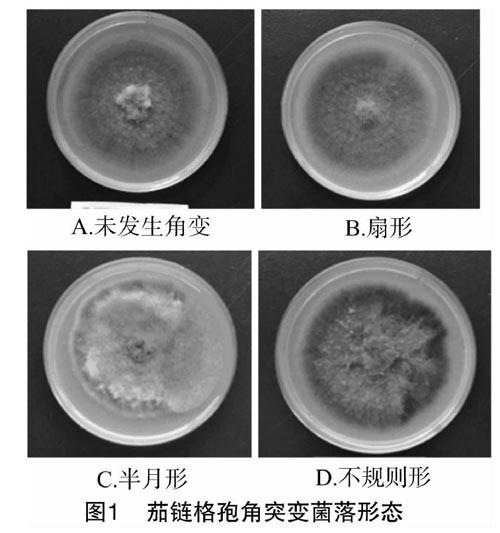
2.1 单孢继代培养对茄链格孢菌落形态的影响
由图1可知,茄链格孢在继代培养过程中出现了扇形、半月形和不规则形等角突变现象。6株亲代菌株在平板培养时有2株发生了角突变现象;每个亲代菌株分别分离到10株F1代菌株,在60株F1单孢后代中共有14株出现了角突变现象,其角突变率为23.33%;通过单孢分离获得的40株F2代菌株中共有21株出现了角突变现象,其角突变率为52.50%。这表明茄链格孢单孢后代随着继代培养代数的增加,其角突变率呈上升趋势。
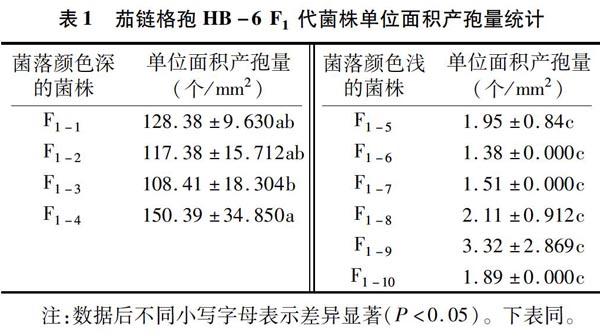
2.2 茄链格孢继代培养菌株菌落颜色与产孢量的相关性
由表1可知,茄链格孢继代培养后代菌落形态与颜色深浅变化明显,且与其单位面积产孢量密切相关,颜色深的单位面积产孢量大,且发生半月形角变的菌株产孢量明显高于未角变菌株(表中未列出)。其中,在HB-6的F1代10株菌株中,菌落颜色深的4株菌株为F1-1、F1-2、F1-3、F1-4,其单位面积产孢量为108~151个/mm2,菌落颜色浅的6株菌株F1-5、F1-6、F1-7、F1-8、F1-9、F1-10单位面积产孢量在1~4个/mm2之间,对菌株HB-6的F1代菌株单位面积产孢量进行方差分析发现,F1-1~F1-4菌株间只有F1-3菌株与F1-4菌株差异显著,其余菌株之间差异不显著,F1-5~F1-10菌株间差异也不显著,但F1-1~F1-4菌株与F1-5~F1-10菌株间差异显著。
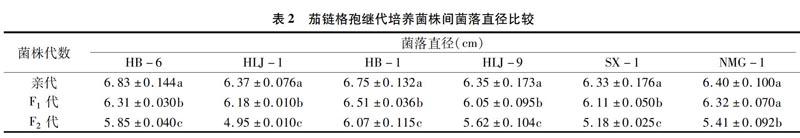
2.3 單孢继代培养对茄链格孢菌株生长的影响
由表2可知,通过十字交叉法测定6株茄链格孢的亲代菌株以及F1、F2代菌株的菌落直径发现,继代培养菌株的菌落直径呈逐代下降趋势。与亲代菌株相比,F1代菌落直径下降1.25%~7.61%,F2代菌株直径下降10.07%~22.29%。其中HLJ-1菌株菌落直径差异最大,与亲代菌株相比,F2代菌落直径下降了22.29%;菌株HB-1下降趋势较缓,F1代菌落直径较亲代下降了3.56%,与F1代相比,F2代菌落直径下降6.76%。对6株茄链格孢的亲代菌株以及F1、F2代菌株菌落直径进行方差分析发现,除菌株NMG-1的亲代和F1代菌落直径间差异不显著外,其余菌株的亲代、F1代、F2代间差异均显著。结果表明,继代培养使菌丝生长速度变慢,相同培养条件下亲代、F1代、F2代的菌落直径呈明显的递减趋势,继代培养抑制了菌丝的生长。
2.4 单孢继代培养对茄链格孢产孢能力的影响
由图2可知,4株茄链格孢菌株随着继代培养,其单位面积产孢量呈下降趋势,且每个菌株的亲代、F1代、F2代之间产孢量差异均显著。与亲代菌株相比,F1代菌株单位面积产孢
量降低12.80%~84.60%。其中,菌株NMG-1的F1代菌株的单位面积产孢量较其余菌株F1代产孢量高,为 27个/mm2,其余菌株产孢量为4~10个/mm2。F2代菌株中,各菌株单位面积产孢量较亲代下降80.11%~96.50%,菌株HLJ-9的F2代菌株单位面积产孢量最低。说明随着继代培养,茄链格孢的产孢能力在不断下降。
3 结论与讨论
继代培养往往会引起微生物菌落形态、颜色、生长和产孢等方面的变化,Rajanikanth等报道,继代培养对白僵菌产孢量的影响很大[10];唐晓庆等研究发现,继代培养会使菌落发生变化[7]。本研究发现,继代培养可以降低茄链格孢的生长速度,同时引起其产孢能力的退化。这与刘海英等关于玉米大斑病菌单孢继代培养的研究结果[3]相符合。
真菌中关于角突变现象的报道已有多个,例如蝉拟青霉菌[11]、禾谷镰刀菌[12]、黄瓜黑星病菌[13]和球孢白僵菌等,徐雍皋等提出,角变现象的产生可能与不正常的细胞核分裂有关[12]。胡海燕等认为,角变现象是异核体分离造成的[14],而唐晓庆等则认为,可能是核内和核外遗传物质共同起作用的结果[15-16]。本研究发现,茄链格孢菌在实验室条件下培养会出现角变现象,F1、F2代菌株出现角变现象的概率分别为 23.33%、52.50%,随着继代培养代数的增加,整体呈上升趋势,这与徐金柱等关于球孢白僵菌角变现象的研究结果[16]相符合。
本研究发现,茄链格孢菌落颜色与产孢量密切相关,菌落颜色深的菌株产孢量显著高于菌落颜色浅的菌株。该结果与张倩倩等关于芸薹生链格孢突变体与野生株相比菌落颜色变浅的研究结果[17]相一致。Lee等通过研究推测单个分生孢子在继代培养过程中可能丢失染色体,茄链格孢单孢继代培养性状的改变可能与该菌染色体丢失引起的变异有关[18]。
参考文献:
[1]Pasche J S,Piche L M,Gudmestad N C. Effect of the F129L mutation in Alternaria solani on fungicides affecting mitochondrial respiration[J]. Plant Disease,2005,81230(3):269-278.
[2]刘 荻,何莉莉,王作乔,等. 继代培养对北虫草菌落形态及子实体产量的影响[J]. 沈阳农业大学学报,2006,37(3):538-541.
[3]刘海英,白亚君,张运峰,等. 继代培养对玉米大斑病菌生长、发育和致病性的影响[J]. 玉米科学,2013,21(1):134-136.
[4]张 慧,吴圣勇,李 娟,等. 不同培养基继代培养球孢白僵菌对西花蓟马毒力和产孢量的影响[J]. 中国农业科学,2016,49(15):2977-2987.
[5]许志斌. 绿僵菌传代菌株的退化[D]. 南昌:南昌大学,2007.
[6]孙召朋. 继代培养对球孢白僵菌毒力及毒力相关酶表达的影响[D]. 长春:吉林大学,2011.[HJ1.72mm]
[7]唐晓庆,黄 勃,李增智. 继代培养对球孢白僵菌抗旱力的影响[J]. 安徽农业大学学报,1996,23(3):289-292.
[8]Meng J W,Zhu W,He M H,et al. High genotype diversity and lack of isolation by distance in the Alternaria solani populations from China[J]. Plant Pathology,2015,64(2):434-441.
[9]刘丽丽. 马铃薯早疫病菌产孢诱导、嘧菌酯敏感性及品种抗病性的研究[D]. 保定:河北农业大学,2013.
[10]Rajanikanth P,Subbaratnam G V,Rahaman S J. Effect of frequency of subculturing of different isolates of beauveria bassiana vuillemin on their biological properties[J]. International Journal of Bio-Resource & Stress Man,2011,2(1):60-65.
[11]胡海燕,刘爱英,邹 晓,等. 一株蝉拟青霉及其角变分离株的初步研究: 第八届海峡两岸菌物学学术研讨会论文集[C].2007:304-309.
[12]徐雍皋,徐敬友,方中达. 禾谷镰刀菌菌落角变的观察[J]. 植物病理学报,1992(1):13-16.
[13]苗则彦,李凤云,赵奎华,等. 黄瓜黑星病菌菌落角变规律研究[J]. 辽宁农业科学,1998(4):17-21.
[14]胡海燕,刘爱英,邹 晓,等. 虫生真菌蝉拟青霉及其角变株的比较研究[J]. 植物保护,2008,34(2):21-25.
[15]唐晓庆,樊美珍,李增智. 球孢白僵菌继代培养中菌落局变现象及环境影响因素的研究[J]. 真菌学报,1996,15(3):188-196.
[16]徐金柱,黄 勃,李增智. 球孢白僵菌单孢子分离株在继代培养过程中菌落局变的遗传分析[J]. 菌物学报,2010,29(2):279-284.
[17]张倩倩,张晓斐,刘伟阳,等. 芸薹生链格孢产孢缺陷突变体的鉴定与表型分析[J]. 基因组学与应用生物学,2017(9):3831-3836.
[18]Lee B N,Adams T H. Overexpression of flbA,an early regulator of Aspergillus asexual sporulation,leads to activation of brlA and premature initiation of development[J]. Molecular Microbiology,1994,14(2):323-334.
