宁夏贺兰山地区狗獾夏季食性研究
2019-08-19王继飞刘振生滕丽微
任 毅 王继飞 高 惠 刘振生,3 滕丽微,3*
(1.东北林业大学野生动物资源学院,哈尔滨,150040;2.宁夏贺兰山国家级自然保护区管理局,银川,750021;3.国家林业和草原局野生动物保护学重点开放实验室,哈尔滨,150040)
狗獾(badger)俗称獾子,是一种体型较小的食肉目(Carnivora)鼬科(Mustelidae)动物,广泛分布于欧亚大陆和北美洲,在我国,除海南和台湾省外,其余各省均有分布[1],为亚洲狗獾(Melesleucurus)。营穴居生活,多白天休息,晨昏活动,具有半冬眠习性。狗獾所产獾油,具有极高的中医药用价值,在治疗烫伤和痔疮等方面有显著效果[2]。目前虽已具备成熟的人工饲养技术,但野外种群常受人兽冲突的影响,近年来各地数量下降迅速[3]。
狗獾食性的相关研究在20世纪初就已进行,目前在世界范围内已经积累了不少的研究成果[4-5],研究区域遍布欧亚大陆,如Murdoch等对蒙古干旱草原地区狗獾食物的研究[6],Balestrieri等对意大利西北部农业河流栖息地的欧洲狗獾(Melesmeles)食性方面的研究[7]等,而国内的相关研究也紧随其后,如李峰等对青海湖盆地狗獾食性的相关研究[4],陈耕对张广才岭地区狗獾食性的对比研究等[8]。经一系列研究发现,狗獾属广食性杂食动物,食物范围涵盖小型哺乳类、鸟类、两栖爬行类、昆虫纲、无脊椎动物、各类植物和果实等,十分广泛。
贺兰山地处宁蒙交界,地理环境独特,狗獾在其中扮演着重要的生态角色,但根据长期外业调查和访问调查情况表明,目前狗獾的数量分布堪忧。其食性方面的研究,对该物种的人工繁育和野外保护方面都尤为重要。因此,本研究采用粪便分析法对贺兰山地区狗獾的夏季食性进行系统研究,以确定其食物种类,通过频率法和生物量比法,确定贺兰山地区狗獾夏季食性的组成,为贺兰山区域狗獾的物种保护提供一定的科学依据。
1 研究地概况
贺兰山位于银川平原和阿拉善高原之间(38°21′—39°22′N,105°44′—106°42′E),主峰海拔3 556.1 m,周围被沙漠、城市和河流(黄河)所阻断。西和北侧为阿拉善戈壁荒漠,东侧为银川平原,是一道处于中国西北地区重要的生态屏障。贺兰山处在典型大陆性气候区域范围内,具有山地气候特征。气候变化大,年均气温-0.8℃,年均降水量420 mm,年均蒸发量2 000 mm。植被有明显的垂直分布规律,随着海拔的升高,植被类型分别为荒漠化草原、山地疏林草原、山地针叶林和亚高山灌丛草甸或高山草甸[9]。代表植物有短花针茅(Stipabreviflora)、黄刺玫(Rosaxanthina)、蒙古扁桃(Amygdalusmongolica)、荒漠锦鸡儿(Caraganaroborovskyi)、折枝绣线菊(Spiraeatomentulosa)、青海云杉(Piceacrassifolia)、灰榆(Ulmusglaucescens)等[10];代表动物有岩羊(Pseudoisnayaur)、金雕(Aquilachrysaetos)、马麝(Moschuschrysogaster)、蓝马鸡(Crossoptilonauritum)、马鹿(Cervuselaphus)、赤狐(Vulpesvulpes)、亚洲狗獾*、草原沙蜥(Phrynocephalusfrontalis)等。目前已知有昆虫纲18目952种,占宁夏昆虫已知种类的41.1%,优势目是鞘翅目(Coleoptera)、鳞翅目(Lepidoptera)、半翅目(Hemiptera)、双翅目(Diptera)和直翅目(Orthoptera),占总科数的62.4%,其中包括螳螂(Mantodea)、草蛉(Chrysopaperla)、步甲类(Carabidae)等对森林有益的天敌昆虫,也有中华真地鳖(Eupolyphagasinensis)、华北蝼蛄(Gryllotalpaunispina)、黄缘龙虱(Cybisterjaponicussharp)等药用昆虫[11]。
2 研究方法
2.1 粪便收集
2018年6—8月,根据贺兰山各个沟段的分布情况和访问调查,确定狗獾的分布范围,于贺兰山马莲口、大水沟管理站下辖各个沟道为基础,制定相应路线,采集狗獾粪样,风干后用保鲜袋存放。研究期间共收集狗獾粪样29份。
2.2 实验步骤
(1)将粪样装入信封袋,用恒温干燥箱在75℃—80℃下干燥24 h,之后称重。
(2)把称重后的样品放入烧杯用肥皂水浸泡12 h,用以去除异味和水溶性物质。
(3)为了加速分离,加入少量的温水或者用解剖针和镊子进行辅助。
(4)待样品充分浸泡后,首先用镊子拣出漂浮在水面的毛发和昆虫残肢等,然后用筛网滤出沉淀物,滤出的溶液倒入烧杯中备用。
(5)反复冲洗沉淀物,将滤出物放于解剖盘内,在水中分拣残余物。
(6)待分拣出的残余物晾干后,烘干称重,装入密封袋中,以备分析。
(7)将之前烧杯中的过滤液先沉淀10 min,然后从大烧杯的底部用宽口移液管移出1.5 mL的次级样本,放入一次性的计数培养皿(培养皿的底部划有均匀分布的1 cm2的方格)中。用苦味酸溶液(用无水乙醇溶解而得)对次级样本进行染色,然后在40倍显微镜下观察蚯蚓的刚毛。随机在培养皿的方格区域中找出10个方格计数蚯蚓的刚毛数,最后计算出1 cm2内蚯蚓刚毛数的平均值。
2.3 实验鉴定
根据分离出的各食物组分,大致可以分为昆虫类、软体动物类、植物类等3大类。昆虫类具体参照《昆虫分类学》[12]、《宁夏贺兰山昆虫》[11]等书目进行辨识,再结合相关专业的专家对存疑残渣进行二次鉴定。但由于残肢细小难辨,仅鉴定至目。软体动物类包括蚯蚓、蜗牛和昆虫幼虫等,消化程度均较高,除蚯蚓可通过刚毛分析,蜗牛可通过残壳称重区分外,其余根据残余情况统归昆虫幼虫,不做细分。植物则多以种子和叶根为主,具体通过《西北农牧交错带常见植物图鉴》[13]、《贺兰山植被》[14]等书目,对比周边采食区植物种类作确认分析。不过除种子外,其他植物碎片多细小破碎,加之有外部沾染因素影响,故不具体区分,通归一类。
此外,还存在一些杂物主要是沙子、碎石及人类生活垃圾等,属于后期沾染,不作分析。
2.4 数据处理
本文主要采用频率法和生物量比法对粪样内容物进行对比分析。每种食物组分在粪样中只要出现,那么其出现频次即算作1,该食物组分的出现频率就是该食物组分在所有粪样中的出现频次与所有食物组分的总出现频次的比值。除蚯蚓外,生物量由粪样辨析出的残留物称重后乘以对应消化系数(昆虫-5[15];果实种子-14[15];其他植物-4[16])即得。蚯蚓生物量利用之前得到的1 cm2内刚毛平均值与生殖环的数量关系(每条蚯蚓只有1个生殖环),估算出蚯蚓的数量,再乘以每只蚯蚓的平均质量2 g(实测数据),便可得到蚯蚓的生物量[8]。对所有粪样采用频率法和生物量比法进行分析,对于所得出的序位结果在SPSS 19.0中进行Spearman等级相关检验。
利用Shannon-Wiener指数H′、均匀性指数E、标准Levins生态位宽度指数Bsta对贺兰山狗獾夏季食物组成的多样性进行对比分析,具体公式如下。
Shannon-Wiener指数公式按:
(1)
均匀性指数公式按:
(2)
标准Levis生态位宽度指数公式按:
(3)
公式中,S为猎物的种类数,Pi为第i种食物组分的生物量比,Bmax为食物组分类型的总数。
3 研究结果
3.1 狗獾夏季食性数据
通过粪样分析法对29份夏季狗獾粪样进行分析,我们共分析出10种食物类型,可分为6大类,按照出现频率多少,依次为昆虫(38.22%)、果实(13.78%)、蚯蚓(12.89%)、昆虫幼虫(12.00%)、其他植物(12.00%)、蜗牛(11.11%),除昆虫外其余类型的出现频率均维持在12%左右,而在昆虫和果实中,作为出现频率最高的鞘翅目(12.89%)和刺玫(Rosasp.,12.89%)果也只是与这一均值持平,而山杏(Armeniacasibirica,0.89%)的出现频率则垫底。从生物量比来看,最大的是蚯蚓(61.83%),其次是果实(29.42%),其中刺玫果(27.37%)贡献了大部分。而昆虫(4.05%)和昆虫幼虫(3.15%)则均在3.5%左右,只有蜗牛(0.62%)和其他植物(0.92%)的生物量比不足1%,包括昆虫里的蜚蠊目(Blattaria,0.78%)、直翅目(0.22%)和膜翅目(Hymenoptera,0.08%)(表1)。夏季狗獾食性的Shannon-Wiener指数H′为1.077,均匀性指数E为0.468,标准Levins生态位宽度指数Bsta为0.131。
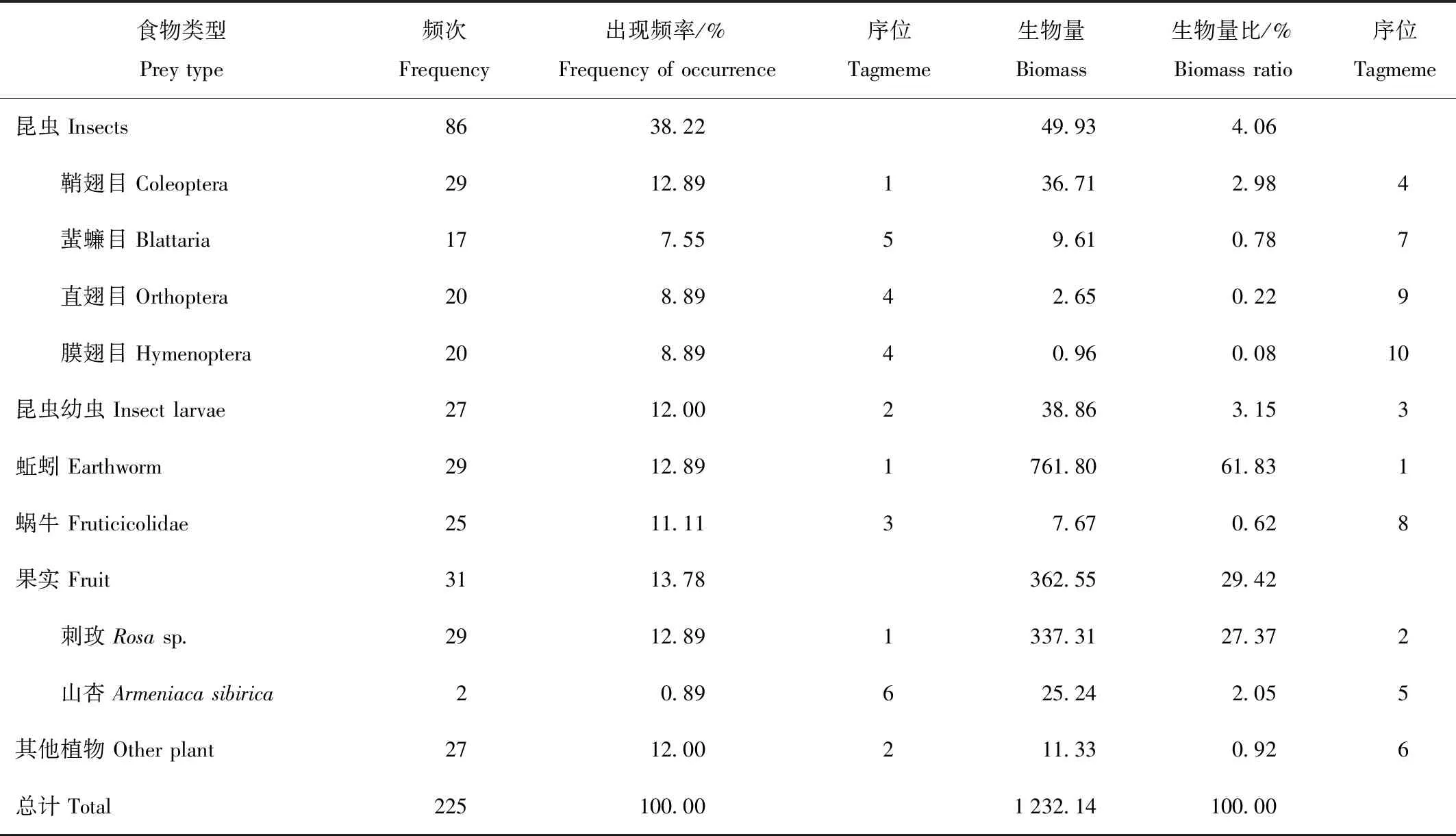
表1 狗獾夏季食物组成比例Tab.1 Diet composition of Meles leucurus in summer
对狗獾食性所得的出现频率与生物量比的序位结果进行Spearman等级相关检验,结果显示,在置信度(双侧)为0.05时,二者没有统计学意义上的显著性相关(P=0.117),这表明这两种方法在对主要食物组成的分析评述上存在差异,因此在两者的结合分析中可以很好地弥补实验的严谨性。
3.2 与其他地区同类食性差异的简要对比
近年来,国内外关于野生狗獾食性的相关研究一直未曾间断[6,8,17-19],以最近的陈耕[8]在张广才岭进行的狗獾食性研究为例,两地夏季食性组成的参数对比如下。
贺兰山食物组分出现频率大小关系为蚯蚓=刺玫果=鞘翅目>昆虫幼虫=其他植物>蜗牛>直翅目=膜翅目>蜚蠊目>山杏。生物量比大小关系为蚯蚓>刺玫果>昆虫幼虫>鞘翅目>山杏>其他植物>蜚蠊目>蜗牛>直翅目>膜翅目。
张广才岭地区食物组分出现频率大小关系为蚯蚓>鞘翅目>半翅目>草树叶>其他植物>蜗牛>膜翅目>昆虫幼虫。生物量比大小关系为蚯蚓>鞘翅目>蜗牛>草树叶>半翅目=其他植物>昆虫幼虫>膜翅目[8]。

表2 贺兰山与张广才岭地区狗獾夏季食性多样性指数、均匀性指数和食物生态位宽度Tab.2 The summer diet composition diversity,evenness index and food niche width of Meles leucurus in Helan Mountain and Zhangguangcai Mountain area
注:因原文中张广才岭地区狗獾夏季食性的指数参数选用的是食物大类数据,为保证对比的准确性和科学性,故而转换用食物小类数据统计张广才岭地区食性指数参数
Note:Because the index parameters of summer feeding habits of badger in the original paper is the food category data,in order to make sure the accuracy and scientific of the comparison,thus converse to use the food subgroup data to statistics the feeding habits’ index parameters of the Zhangguangcai Mountain area
4 讨论
粪样分析法虽具有非损伤性和不干扰动物正常活动的优点,但是也存在诸如部分残余物无法辨认[20],归属鉴定难等问题,而且容易忽视一些完全被消化的食物组分[21]。在本次研究中,我们也遇到了相同的问题,如对蚯蚓的鉴定只能通过刚毛的存在及生殖环比例来推算,而昆虫及幼虫也因消化的问题变得细小破碎,致使造成一部分食物类型无法做到更详尽的鉴定。
作为粪便分析中常用的两个方法,频率法简单易行,能很好地反映出动物捕食猎物的种类及次数,生物量比法则可以完整地体现捕食者对该类食物项目捕食量的多与少[22],因而通过两者能够最大程度上体现出各食物类型在动物食谱中的相对重要性。
一直以来,狗獾都被认为是杂食性的广食捕食者,这一点在现有诸多文献中均有体现[6-8,19]。但在早期,部分研究者在一些研究地区发现,那里的狗獾几乎只吃蚯蚓,因此狗獾曾一度被认为是蚯蚓的专食者[17],而地中海地区的狗獾却很少摄食蚯蚓[18]。后经研究发现,狗獾主要是根据食物的可获得性程度来利用资源,同时地理环境的差异也会导致食物资源的可获得性存在差异,在低纬度地区,环境中的爬行动物和鼠虫等食物资源更丰富一些,而在相对湿润的高纬度地区,蚯蚓量则比较丰富[4]。
4.1 狗獾夏季食性
对于宁夏贺兰山地区的狗獾来说,其夏季的食物组成主要来自于蚯蚓和刺玫果,出现频率均呈现最高,而两者的生物量占比达89%,蚯蚓就占近七成。对于蚯蚓的摄食,与其他地区狗獾食性研究的特点一致,同时也符合作为穴居动物,蚯蚓的获取容易度和可遇见性要远高于其他种类的昆虫及幼虫的推断。而遍布贺兰山地区的蔷薇科(Rosaceae)刺玫类植物,以其可口且营养的果实深受狗獾喜爱,广泛的分布和低矮的植株也是狗獾摄取的重要因素。

图1 贺兰山与张广才岭地区狗獾夏季食性组成对比Fig.1 Comparison of summer dietary composition of Meles leucurus between Helan Mountain and Zhangguangcai Mountain area 注:因原文中张广才岭地区狗獾夏季食性出现频率计算方式采用的是该食物组分在所有粪样中的出现频次与粪样总数的比值,为保证对比的准确性和科学性,故而转换用与所有食物组分的总出现频次的比值。同时原文中的蠕虫即为本文中的昆虫幼虫 Note:Because the calculating method of summer feeding frequency of badger in Zhangguangcai Mountain area in the original paper used is ratio of the frequency of the food component in all feces to the total fecal samples,in order to make sure the accuracy and scientific of comparison,thus converse to the ratio of the total occurrence frequency of all food components.At the same time,the worm in the original paper is the insect larva of this paper
同时从研究中可以看出,昆虫类的出现频率很高,但生物量比却很低,一方面可能是消化系数值较小的缘故[23],另一方面可能因为昆虫容易被狗獾消化得更彻底,而且细小的食物残渣通过筛网流出的较多。而昆虫中,占比最多的是鞘翅目,其变态发育前后营土栖生活的习性和欠佳的飞行迁移能力使其相较于其他昆虫更易被捕食。该目在贺兰山属优势目,种类和数量众多,且其中多数种类可危害农林作物[11]。对比之前的研究,食物中蜚蠊目和直翅目的发现也增加了狗獾食性研究的数据资料。
本研究中将蜗牛和昆虫幼虫各分为一大类,主要是因为其在狗獾粪便残余中的特征与其他昆虫明显不同,虽然在出现频率上,二者处在食物组分中相当重要的地位,但从生物量比来说,两者并不是狗獾食物的主要来源,除去可遇见性的原因外,也反映出狗獾对其偏好程度维持在较低状态。
对于粪便中发现的其他植物,在除去外部沾染的因素下,我们发现部分植物根叶具啃咬痕迹,因而可以确定是狗獾主动采食的结果,其原因很大程度上可能是植物类食物的粗纤维能够帮助狗獾顺利排泄难以消化的粪便[24]。
意外的是,夏季的粪便中并未发现鸟类和两栖爬行类乃至哺乳类的痕迹残渣,据推测,可能与狩猎难度和能量利用存在一定关系,相较于蚯蚓昆虫和植物果实,鸟类、两爬和哺乳类的获取难度要高出数倍,同时所付出的能量和获得的收益也难成正比。
综上所述,贺兰山的夏季,因雨水充沛,蚯蚓和昆虫大量繁殖,果实挂果,狗獾无须花费更多的力气去找寻其他食物,这个时候主要采食蚯蚓、昆虫和果实。因此在夏季,作为能量高易取食的代表食物蚯蚓,不论是在出现频率还是在生物量比中,都占狗獾食物组成的绝大部分,而分布广泛但能量相对较低的果实为狗獾的采食提供了有效的补充。同时其他昆虫及幼虫因遇见性和能量的关系,出现频率很高,生物量比却远低于蚯蚓,以上研究结果再次印证了狗獾杂食性广食者的食性定位。
4.2 与其他地区同类食性的差异
根据图1可以看出,一方面两地狗獾在夏季食性组成中均具备各自特有的食物组分,如昆虫中的半翅目、蜚蠊目、直翅目,果实中的刺玫果、山杏等(草树叶可能属沾染杂质,在原文数据分析中并未使用,为确保准确性,这里也不做考虑),其中不乏像半翅目和刺玫果一样,在各自食性中占据相当地位的食物项目,充分反映出狗獾在不同自然环境中食性的适应能力。另一方面假如去除两地狗獾的特异性食物组分不看,在其余共有的食物组分里,无论是出现频率还是生物量比,所占比例和重要性均十分相近。如蚯蚓都是两地狗獾的主要食物,在出现频率和生物量比上均占据关键地位。在其余昆虫中,也都是鞘翅目占比更重,均具有较高的出现频率和较低的生物量比。
根据表2所示,在Shannon-Wiener指数H′和均匀性指数E上,张广才岭地区呈现出一定量的偏高,而贺兰山地区则在食物生态位宽度指数Bsta上略高一筹,但相较于同域分布的其他动物(赤狐Vulpesvulpes)来说[24],依旧较窄。在排除实验误差外,其实两地参数的差值并不大。在对两地生物量比数据进行融合后,通过独立性T检验分析,二者不存在显著性差异(P=0.965),即两地狗獾在夏季食性上差异不显著。但从地理环境来看,两地均位于北纬40°以上,其中宁夏贺兰山气候干燥,岩石遍布,土层贫瘠,植被稀少,而张广才岭地区气候湿润,土层肥沃,植被密布,气候和环境差异巨大,即便在食物选择相对丰富,生物量较多的夏季,也未影响两地狗獾在食物结构和选择上的相似性。本研究结果可为动物食性与环境间相互作用的研究提供后续探究的思路。
*注:目前研究一般将狗獾分为欧洲狗獾和亚洲狗獾两个种[25],本研究认为宁夏贺兰山地区的狗獾属于亚洲狗獾。
