氮素形态对盐胁迫下辣椒幼苗生长及光合特性的影响
2019-08-15刘会芳韩宏伟庄红梅
刘会芳,韩宏伟,庄红梅,王 强,宋 羽,王 浩
(1.新疆农业科学院园艺作物研究所,乌鲁木齐830091;2. 新疆农业科学院农作物品种资源研究所,乌鲁木齐830091)
0 引 言
【研究意义】氮素作为酶的主要成分广泛参与到植物光合作用的各个环节,氮素形态及供氮水平直接影响植株的光合效率,进而影响产量和品质[1]。在设施蔬菜栽培过程中,化肥的不合理施用等因素,造成盐分在土壤中积聚,破坏了养分平衡,引起土壤的次生盐渍化,影响了作物氮素的吸收和同化[2]。因此,确定设施环境内蔬菜栽培过程中适宜的供氮水平,有利于提高植物的光合能力,进而达到增加产量和提升品质的效果。【前人研究进展】诸多研究表明,植物体吸收利用的氮素营养存在铵态氮(NH4+)、硝态氮(NO3-)和小分子有机氮3种形态,由于它们具有不同的物理化学性质,在植物体吸收后通常会使植物体显示出明显的生长差异[3]。氮素营养也可以缓解盐胁迫对植物造成的伤害[4-5]。Nathawat等[6]的研究表明,与铵态氮相比,添加硝态氮,芥菜耐盐性更强,与油菜上[7]的结果相似,与大麦上[8]的研究结果相反。【本研究切入点】在设施辣椒中的研究还鲜有报道。研究辣椒幼苗在盐胁迫条件下对不同氮素形态的响应。【拟解决的关键问题】以辣椒幼苗为研究对象,采用营养液培养,通过供应等氮量的硝态氮、铵态氮和有机态氮条件下,研究不同氮素形态对盐胁迫下辣椒幼苗壮苗指数、根冠比、光合气体交换参数及叶绿素荧光参数等的影响。为设施辣椒科学施肥及盐渍化土壤的利用提供理论依据。
1 材料与方法
1.1 材 料
试验于2018年8月在新疆农业科学院安宁渠试验站日光温室内进行,供试辣椒品种为新椒27号。采用穴盘基质育苗,待幼苗长至四叶一心,转入室内光照培养架,置于不同供氮形态下的营养液中。整个水培系统培养箱大小一致,均为450 mm×320 mm×155 mm,每个培养箱装10 L营养液,每个培养箱定植30株辣椒幼苗,每个处理3个培养箱。整个系统用增氧泵提供氧气,每天24 h不间断供氧。室内环境控制标准:温度控制在白天25℃,晚上18℃,空气湿度40%,光周期为白天14 h/夜晚10 h。
辣椒幼苗适应生长7 d后,加入盐胁迫处理(100 mmol/L NaCl)。不同氮素形态分别为硝态氮(硝酸钾),铵态氮(硫酸铵)和有机态氮(甘氨酸),3种氮素供应形式将改良霍格兰氏营养液中氮元素浓度设为2 mmol/L,调节pH至6.00±0.05。试验共设6个处理:硝态氮营养液(A1)、硝态氮营养液+100 mmol/L NaCl胁迫(A1T),铵态氮营养液(A2)、铵态氮营养液+100 mmol/L NaCl胁迫(A2T),有机态氮营养液(A3)、有机态氮营养液+100 mmol/L NaCl胁迫(A3T)。每个处理3个培养箱,共18个培养箱。试验过程中,每3 d更换一次营养液,每天调节各处理营养液pH至6.00±0.05,直至试验结束。
1.2 方 法
于盐胁迫第3 d,每个重复随机取出6株植株,分别测定幼苗株高、茎粗,并将辣椒植株分为地上部和地下部两部分,将收集的地上部茎、叶和地下部根系用去离子水洗干净,并用吸水纸吸干剩余水分,放入信封,105℃杀青10 min,80℃烘干至恒重,称重,计算壮苗指数和根冠比。壮苗指数=茎粗/株高×全株干质量,根冠比=地下部干质量/地上部干质量。
光合气体交换参数测定:于 11:00用 Li-6 400 便携式光合作用测定仪(Li-COR,USA),设定光照强度为800 umol(m2·s),测定幼苗倒三叶的净光合速率(Pn)、气孔导度(Gs)、胞间 CO2浓度(Ci)和蒸腾速率(Tr)。并计算气孔限制值(Ls):Ls= 1-Ci/Ca。其中,Ca是空气中 CO2浓度。叶片水分利用效率(WUE):WUE=Pn/Tr。羧化效率(CE):CE=Pn/Ci。每个处理测3次重复,每个重复测3片叶子。
叶绿素荧光参数测定:利用PAM—2 500便携式叶绿素荧光仪(Walz,Germany),选取正常辣椒幼苗植株的倒三叶,将叶片暗适应30 min,,随后测定初始荧光(F0)、最大荧光(Fm)、最大光化学效率(Fv/Fm),随后在自然光下适应30 min,测得光下最小荧光产量(Fs)、实时荧光产量(F’)、光下最大荧光产量(Fm’)、实际光化学效率(ΦPSⅡ),并由此计算光化学猝灭系数(qP)=(Fm’-F’)/(Fm’-F0’),其中F0’=1/(1/F0-1/Fm+1/Fm’),非光化学猝灭系数(NPQ)=Fm/Fm’-1
1.3 数据处理
采用Excel软件对数据进行整理,采用SPSS16.0软件对各处理间数据进行显著性差异分析,作图采用Excel软件。
2 结果与分析
2.1 不同氮素形态下盐胁迫对辣椒幼苗生长的影响
研究表明,非盐胁迫条件下,不同氮素形态以铵态氮处理辣椒幼苗壮苗指数最大,显著大于有机态氮,二者又显著大于硝态氮。盐胁迫后,与A1相比,A1T处理壮苗指数显著增加,增加幅度达到47.21%,与A2相比,A2T处理壮苗指数显著降低,降低幅度为17.58%,与A3相比,A3T处理壮苗指数增加了4.21%,未达到显著差异水平。A1T、A2T、A3T三者相比,A1T壮苗指数显著大于A3T,显著大于A2T。盐胁迫对硝态氮处理影响最大。图1
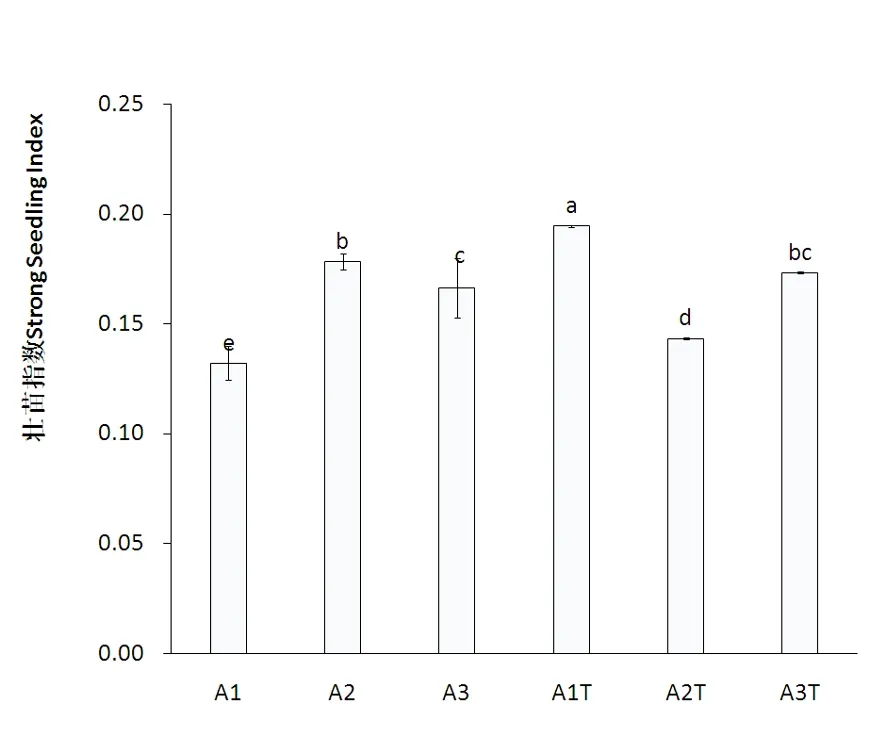
注:图中小写字母表示处理间差异显著性(P<0.05)。下同
Note:The different letters mean significant difference atP<0.05.The same as below
图1 氮素形态对盐胁迫下辣椒幼苗壮苗指数变化
Fig.1 Effects of different N forms on Strong seedling index of the pepper under salt stress
研究表明,非盐胁迫条件下,不同氮素形态以铵态氮处理辣椒幼苗根冠比最大,显著大于硝态氮和有机态氮处理,A1和A3处理之间无显著差异。盐胁迫后,分别与A1和A3相比,A1T和A3T处理根冠比显著增加,增加幅度达到22.68%、41.42%,与A2相比,A2T处理根冠比降低6.12%,未达到显著差异水平。A1T、A2T、A3T三者相比,A3T处理根冠比显著大于A1T和A2T处理,A1T和A2T两处理之间未达到显著差异水平。图2

图2 氮素形态对盐胁迫下辣椒幼苗根冠比变化
Fig.2 Effects of different N forms on Root-shoot ratio of the pepper under salt stress
2.2 不同氮素形态下盐胁迫对辣椒幼苗叶片光合气体交换参数的影响
研究表明,非盐胁迫条件下,不同氮素形态以A1处理的辣椒幼苗叶片的Pn、Cs、Ci和Tr最大,显著高于A2、A3处理。A2处理的Pn、Cs和Tr显著高于A3处理,Ci则表现为A2和A3处理无显著差异。三种氮素形态以硝态氮处理辣椒幼苗的光合作用最强,铵态氮和有机态氮则一定程度的抑制了辣椒幼苗的光合作用,且A2和A3处理Ci无明显差异说明有机态氮处理的光合抑制作用不仅表现为气孔限制。盐胁迫后,与A1相比,A1T处理的光合气体交换参数均有所降低,Pn、Gs和Tr显著高于其它胁迫处理。与A2相比,A2T处理的光合气体交换参数也表现为降低的趋势,Pn、Cs和Tr显著高于A3T处理,Ci则表现为A1T和A2T处理无显著差异,且二者均显著大于A3T处理。与A3相比,A3T处理的Pn则显著增加了21.77%,Gs和Tr与盐胁迫前无明显差异,Ci显著降低了15.12%。盐胁迫抑制了A1和A2两种氮素处理下辣椒幼苗的光合作用,却一定程度的刺激了有机态氮处理下的辣椒幼苗的光合作用。图3
2.3 不同氮素形态下盐胁迫对辣椒幼苗叶片气孔限制值、水分利用效率和羧化效率的影响
研究表明,非盐胁迫条件下,不同氮素形态以A3处理辣椒幼苗叶片的气孔限制值最大,显著高于A2、A3处理,A2处理显著大于A1处理。盐胁迫后,分别与A1、A2、A3相比,A1T、A2T、A3T各处理辣椒幼苗的叶片气孔限制值均显著增加,增加幅度分别为16.86%、3.97%、8.73%。A1T、A2T、A3T三者相比,A3T处理的辣椒幼苗叶片气孔限制值显著大于A2T处理,二者显著大于A1T处理。不同氮素形态下,铵态氮和有机态氮均表现出了一定的气孔限制,盐胁迫后气孔限制均有所增强,以有机态氮增加最大。图4

图3 氮素形态对盐胁迫下辣椒幼苗叶片光合气体交换参数变化
Fig.3 Effects of different N forms on gas exchange parametersof the pepper leaves under salt stress

图4 氮素形态对盐胁迫下辣椒幼苗叶片气孔限制值变化
Fig.4 Effects of different N forms onstomatal limitation of the pepper leaves under salt stress
研究表明,非盐胁迫条件下,不同氮素形态以A1处理辣椒幼苗的叶片水分利用效率最小,显著低于A2、A3处理,A2和A3处理之间无明显差异。盐胁迫后,分别与A1、A2、A3相比,A1T、A2T、A3T各处理的叶片水分利用效率均有所增加,增加幅度分别为:9.13%、2.62%、10.48%,其中A1T和A3T达到显著差异水平。A1T、A2T、A3T三者相比,A3T处理的辣椒幼苗叶片水分利用率显著大于A2T处理,二者显著大于A1T处理。图5

图5 氮素形态对盐胁迫下辣椒幼苗叶片水分利用率变化
Fig.5 Effects of different N forms onwater use efficiencyof the pepper leaves under salt stress
研究表明,非盐胁迫条件下,不同氮素形态以A1处理辣椒幼苗叶片的羧化效率最大,显著大于A2、A3处理,A2处理显著大于A3处理。盐胁迫后,各处理辣椒幼苗叶片的羧化效率表现不一,与A1相比,A1T显著降低,降低幅度为20.05%,与A2相比,A2T也表现为降低,降低幅度为13.13%,未达到显著差异水平,与A3相比,A3T则显著升高,升高幅度为53.26%。A1T、A2T、A3T三者相比,A1T和A2T两处理之间无明显差异,二者均显著大于A3T处理。不同氮素形态下,硝态氮能够使辣椒幼苗叶片保持较高的羧化效率,盐胁迫后虽显著下降,仍是最高。图6

图6 氮素形态对盐胁迫下辣椒幼苗叶片羧化效率变化
Fig.6 Effects of different N forms oncarboxylation efficiencyof the pepper leaves under salt stress
2.4 不同氮素形态下盐胁迫对辣椒幼苗叶绿素荧光参数的影响
研究表明,初始荧光(F0)和最大荧光(Fm)表现有所不同。非盐胁迫条件下,不同氮素形态下A1和A2两处理辣椒幼苗叶片的F0显著低于A3处理,且A1和A2两处理之间无显著差异。Fm则表现为A1处理显著大于A2处理,两者显著大于A3处理。盐胁迫后,与A1相比,A1T处理的F0显著增加,Fm显著降低;与A2相比,A2T处理的F0和Fm均显著增加,与A3相比,A3T处理的F0显著降低,Fm显著增加。A1T、A2T、A3T三者相比,F0表现为A1T处理显著大于A2T和A3T两处理,Fm表现为A1T和A2T两处理显著大于A3T处理。图7
研究表明,非盐胁迫条件下,不同氮素形态A1处理辣椒幼苗叶片的实际光化学效率(ΦPSⅡ)大于A2处理,未达到显著差异水平,最大光化学效率(Fv/Fm)显著大于A2处理,A2处理的ΦPSⅡ和Fv/Fm均显著大于A3处理。盐胁迫后,与A1相比,A1T的ΦPSⅡ增加,Fv/Fm降低,均未达到显著差异水平,分别与A2和A3相比,A2T和A3T两处理均表现为ΦPSⅡ显著降低,Fv/Fm无明显变化。A1T、A2T、A3T三者相比,A1T处理的ΦPSⅡ显著大于A2T处理,Fv/Fm无明显差异,两处理均显著大于A3T处理。图8

图7 氮素形态对盐胁迫下辣椒幼苗叶片初始荧光和最大荧光变化
Fig.7 Effects of different N forms oninitia and maximum fluorescenceof the pepper leaves under salt stress

图8 氮素形态对盐胁迫下辣椒幼苗叶片实际光化学效率和最大光化学效率变化
Fig.8 Effects of different N forms onmaximum actual photochemical efficiencyof the pepper leaves under salt stress
研究表明,非盐胁迫条件下,不同氮素形态A1、A2、A3处理辣椒幼苗叶片的光化学猝灭(qP)均无明显差异,非光化学猝灭(NPQ)则表现为A1处理显著大于A2处理,二者均显著大于A3处理。盐胁迫后,与A1相比,A1T处理的qP显著增加,NPQ显著降低,与A2相比,A2T处理的qP无明显变化,NPQ显著增加,与A3相比,A3T处理的qP显著降低,NPQ显著增加。A1T、A2T、A3T三者相比,qP表现为A1T处理显著大于A2T处理,二者显著大于A3T处理,NPQ表现为A1T处理显著小于A2T和A3T处理,后两者之间无显著差异。图9

图9 氮素形态对盐胁迫下辣椒幼苗叶片光化学猝灭和非光化学猝灭变化
Fig.9 Effects of different N forms onphotochemical and non-photochemical chloropyll fluorescence quenching of the pepper leaves under salt stress
3 讨 论
植物的生长发育受不同形态氮素的调控,不同形态的氮素营养可以通过影响植物的诸多方面影响植物的生长发育。壮苗指数和根冠比是评价幼苗质量的重要指标。研究结果表明,铵态氮条件下辣椒幼苗的壮苗指数和根冠比最大,辣椒作为喜硝作物,硝态氮供应条件下,壮苗指数和根冠比小于铵态氮可能是由于,辣椒幼苗在氮素供应充足时,大部分氮素与光合产物用于枝叶生长,供应根部的数量相对较少,所以根冠比降低,而铵态氮供应条件下,辣椒幼苗吸收的氮素首先满足根的生长,运到地上部的就减少,所以根冠比增大。盐胁迫后,硝态氮处理的壮苗指数显著增加,说明盐胁迫刺激了辣椒幼苗的生长。
光合气体交换参数等指标主要用于直观性反映植物光合作用情况。试验结果表明,非盐胁迫条件下,硝态氮处理辣椒幼苗较铵态氮和有机态氮处理气孔限制值最小,Gs、Ci、Pn和CE最大,说明硝态氮处理能够使辣椒幼苗叶片保持较强的气孔活性,维持较高的叶片光合碳同化能力,有利于促进光合产物的积累,促进其生物量的积累;硝态氮叶片WUE明显小于铵态氮和有机态氮,这可能由于叶片高氮比低氮条件下植物内部水分利用效率减小, 其原因是光合速率比蒸腾速率减小的要大,这在其后的辣椒幼苗不同器官氮含量测定试验中也得到了证明。曹生奎等[8]研究表明,在较干旱的生境里,具有较高的WUE,使用较保守的水分利用策略以允许植物生长、生存和维持。而在较湿润环境里,较低的WUE和挥霍的水分利用模式能使植物获得较高的生产力。盐胁迫后,硝态氮较胁迫前对应氮素处理光合速率和羧化效率下降,Gs和Ci的降低以及Ls的增大说明其降低主要是由于气孔限制因素,叶片WUE增加以及Tr降低说明盐胁迫条件下辣椒幼苗通过降低蒸腾速率来提高叶片水分利用率。铵态氮较胁迫前对应氮素处理变化趋势与硝态氮相似,其叶片水分利用率和羧化效率无显著变化,说明辣椒叶片净光合速率的降低并不是由于碳同化第一步羧化作用的降低导致,具体原因还有待进一步试验证实。有机态氮较胁迫前对应氮素处理净光合速率显著增加,Ls、Gs和Ci的变化说明其增加是由非气孔因素导致,Tr无明显变化加之WUE的显著增加暗示盐胁迫改变了辣椒幼苗的水分利用模式,获得较高的光合速率和羧化效率。
叶绿素荧光动力学参数主要用于表征叶片光合作用过程中光系统对光能的吸收、传递、耗散和分配的特征,能有效的反映出叶片内部光合元件的运转情况[9]。试验中,非盐胁迫条件下,硝态氮处理辣椒幼苗叶片F0较有机态氮处理低,说明硝态氮处理更有利于维持PSⅡ反应中心的稳定性;Fm、Fv/Fm、ΦPSⅡ高于铵态氮和有机态氮处理,说明硝态氮有利于提高辣椒叶片PSⅡ反应中心的光化学活性,从而增强其对光能的利用效率;NPQ高于铵态氮和有机态氮处理,说明辣椒幼苗能够把过剩的光能转化为热能耗散,从而保护自身的光合生理功能。盐胁迫后,硝态氮较胁迫前对应氮素处理F0和qP显著增加、Fm和NPQ显著降低,说明辣椒幼苗叶片的PSⅡ的稳定性被破坏,放氧复合体(OEC)失活,但PSⅡ反应中心氧化态QA 的比例增加,进而促进了光合活性增加[10]。铵态氮较胁迫前对应氮素处理F0、Fm和NPQ显著增加、ΦPSⅡ显著降低,说明辣椒幼苗叶片的放氧复合体(OEC)活性增强但其PSⅡ的稳定性可能遭到被破坏,且其将过剩光能耗散为热的能力降低,自我保护能力下降,从而导致其实际光化学效率降低。有机态氮较胁迫前对应氮素处理Fm和NPQ显著增加、F0、ΦPSⅡ和qP显著降低,说明辣椒幼苗叶片的PSⅡ反应中心氧化态QA 的比例减小,放氧复合体(OEC)活性增强,且其耗散过剩光能的能力增强,但这同样也降低了光能用于光化学反应的比例,这会直接导致辣椒叶片同化力( ATP 和NADPH) 的供应,进而抑制其光合碳同化能力[11]。这与其他喜硝氮植物在以其它氮素(如铵态氮)为单一氮源时,植物光合作用受到抑制,光能利用能力降低,从而对植物造成毒害的研究结果相似[12]。
4 结 论
三种不同的氮素供应形态相比,单一硝态氮和铵态氮源的供应均能够使辣椒幼苗叶片光系统保持其稳定性和功能性,从而有较强的光合能力和光能利用能力。辣椒幼苗的光合气体交换参数(Pn、Gs、Ci、Tr)均显著下降,气孔限制值(Ls)显著增加,说明二者光合速率下降的主要原因是气孔限制。F0和qP的显著增加说明光系统Ⅱ稳定性可能遭到破坏,但硝态氮处理的辣椒幼苗叶片能够通过增加PSⅡ反应中心氧化态QA 的比例使光合活性增加,最终增加了辣椒幼苗的壮苗指数47.21%和根冠比22.68%。铵态氮处理虽然能够增强放氧复合体(OEC)活性,但其耗散过剩光能的能力降低导致自我保护能力下降,从而导致实际光化学效率(ΦPSⅡ)显著降低,最终导致壮苗指数显著降低17.58%。其中硝态氮供应效果明显更好。而有机态氮的供应则对辣椒幼苗造成了一定程度的毒害,表现在辣椒幼苗Pn和Ls显著增加,Ci显著降低,辣椒幼苗能够减少叶片蒸腾增加叶片水分利用率,使叶片保持较高的羧化效率,增加了辣椒幼苗叶片的净光合速率,但其Fm和NPQ显著增加、F0、ΦPSⅡ和qP显著降低,PSⅡ反应中心虽放氧复合体活性增强,但氧化态QA的比例减小,加上光能的过度耗散使叶片同化力降低,但其壮苗指数和根冠比的增加结合盐胁迫前试验结果则说明单一有机态氮供应使辣椒幼苗受到毒害。三种不同的氮素供应形态相比,硝态氮供应条件下辣椒幼苗叶片光系统其稳定性和功能性保持最好,光合能力和光能利用能力最强,硝态氮作为设施辣椒施肥及盐渍化土壤的利用氮源。
