藏猪EGLN1基因exon 4及部分内含子变异研究
2019-08-12李卓航寸博文文禹粱赵生国
李卓航,寸博文,文禹粱,赵生国,蔡 原
(甘肃农业大学动物科学技术学院,甘肃兰州730070)
藏猪(Sus scrofa)等青藏高原土著动物形成了一种可以研究人类与动物适应低氧的“群体优势”[1],最终表现在动物组织结构特征和基因差异表达等方面[2-3]。对甘孜、迪庆、合作及西藏藏猪促红细胞生成素(EPO)基因的研究表明,EPO 具有独特的单核苷酸及氨基酸变异[4];在基因组水平上筛选出了藏猪更多低氧适应相关基因和SNPs(单核苷酸多态性)[5]。有研究表明,脯氨酰羟化酶蛋白(EGLN1)基因与藏族人血液中血红蛋白浓度关系密切[2],但针对猪的低氧适应相关研究较少。EGLN1 基因又被称为PHD2,是一个关键的氧传感基因[6],也是最早发现与低氧有关的基因。近年来研究表明,脯氨酸羟化酶(PHD)是脯氨酸羟化酶家族的一个成员,其在缺氧诱导因子的羟基化调解中起着重要作用,被认为是细胞的“氧感受器”[7-8]。本文对藏猪EGLN1 基因exon 4 及部分核苷酸序列进行测序分析,以期为探索和揭示藏猪低氧适应的分子学机制提供参考,也对挖掘藏猪优良种质资源、维系青藏高原生态系统稳定具有理论意义。
1 材料与方法
1.1 实验动物 本研究分别从不同海拔地区采集了4 个藏猪群体(西藏藏猪、合作藏猪、甘孜藏猪和迪庆藏猪)、八眉猪、烟台黑猪及2 个引入种猪(长白猪和大白猪)共35 头猪的组织样品(表1)。
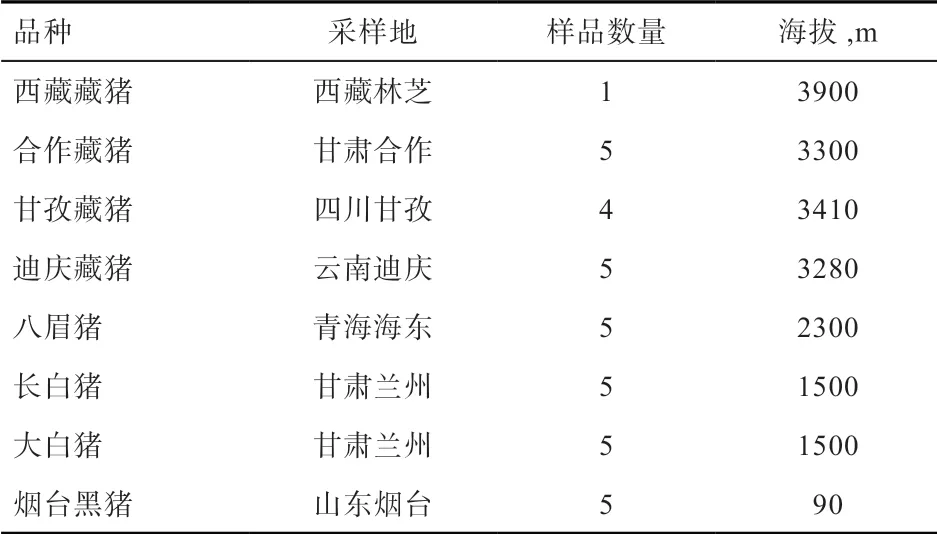
表1 样品信息
1.2 实验方法 采用常规酚- 氯仿抽提法提取基因组DNA。根据杜洛克猪EGLN1 基因全序列(登录号NW_001885473)设计引物(表2),由上海生工生物工程有限公司合成。PCR 扩增采用50 μL 体系进行扩增,反应条件:94℃预变性3 min,94℃变性30 s、56.5℃退火30 s、72℃延伸30 s、共30 次循环,72℃延伸10 min,4℃保存。扩增产物经2.0% 琼脂糖凝胶、150 V 电压下电泳检测20 min,将条带清晰明亮目的产物送至北京六合华大基因科技股份有限公司进行测序。
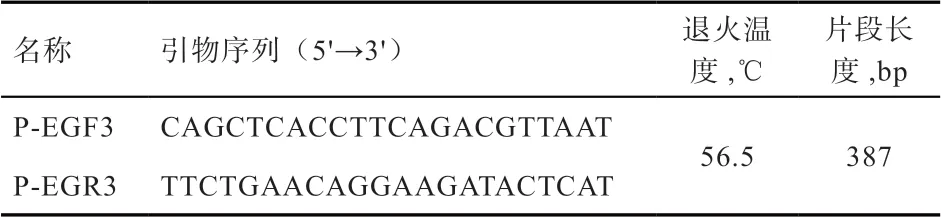
表2 引物信息
1.3 统计分析 用Chromas Version 2.33 对原始序列进行编辑,利用MEGA5.0 及ClustalX 软件进行同源序列比对分析,运用PopGen32 软件计算基因型频率和等位基因频率。
2 结 果
2.1 EGLN1 基因exon 4 等位基因检测 EGLN1 基因exon 4 核苷酸序列变异位点分析采用Chromas Version 2.33 软件对测序结果进行编辑,获得全长为333 bp 核苷酸序列,位于已发表的杜洛克猪EGLN1 基因全序列(NW_001885473) 的48 090~48 422 bp 处。 采 用MEGA7.0 软件对西藏藏猪、合作藏猪、甘孜藏猪、迪庆藏猪、八眉猪、长白猪、大白猪、烟台黑猪8 个群体共35 条核苷酸序列分析表明,EGLN1 基因exon 4 核苷酸序列在266 bp 处发生了C→T 的颠换,4 个藏猪群体和八眉猪在第266 碱基处均为C,而3 个选育猪种(长白猪、大白猪和烟台黑)在该处存在C/T 杂合及TT 纯合现象(图1)。
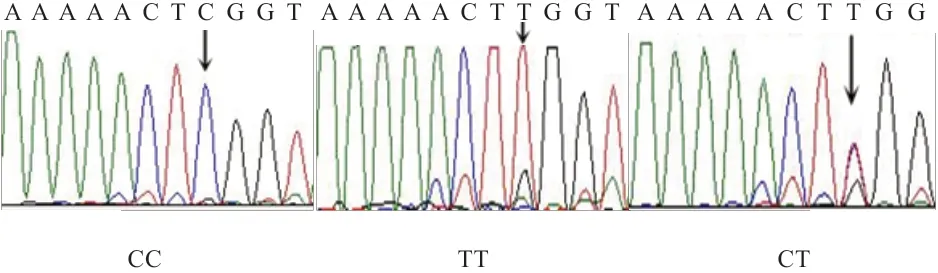
图1 EGLN1 基因exon 4 PCR 测序电泳图
2.2 EGLN1 基因exon 4 基因型分析 根据EGLN1 基因exon 4 上266 bp 处发生的颠换,界定出CC、CT 和TT 3 种基因型。对8 个猪群体的基因型分析发现(表3),高原猪种4 个藏猪群体和八眉猪群体只存在CC 基因型,而3 个选育猪种(长白猪、大白猪、烟台黑猪)共15个个体存在3 种基因型,其中CC 型有5 个、CT 型有6 个、TT 型有4 个。基因型频率分析表明,CC 基因型在藏猪和八眉猪中具有绝对优势,频率均为1,而3 个选育猪种中3 种基因型都存在,频率介于0.20~0.60。就基因频率而言,藏猪和八眉猪只有等位基因C,而等位基因T 在选育猪种中的频率有0.40。
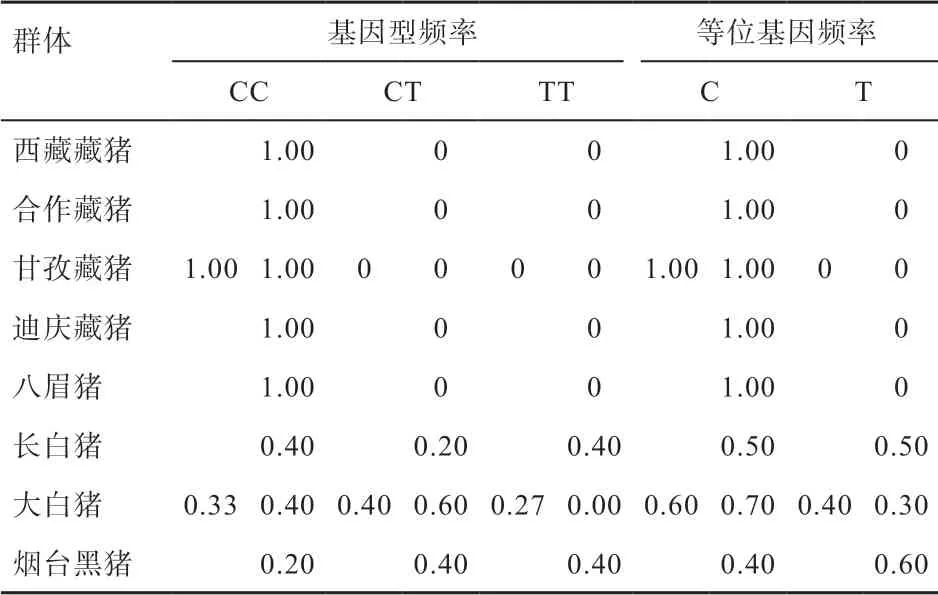
表3 EGLN1 基因exon 4 基因型频率和基因频率
3 讨 论
近年来研究表明,EGLN1 基因在高海拔低氧环境适应的人群中存在很强的正选择,它是欧亚人群(包括藏族)和安第斯人群共同的与高海拔低氧环境适应相关的基因[9-11]。EGLN1 受选择的范围很宽广,其基因多态性与血红蛋白含量较低相关,而较低的血红蛋白含量正是藏族适应低氧环境的生理反应的原因之一[12]。本研究对4 个藏猪群体(西藏藏猪、甘孜藏猪、迪庆藏猪、合作藏猪)、八眉猪和选育的大白猪、长白猪、烟台黑猪EGLN1 基因分析发现,在第4 外显子的266 bp 处发生了碱基C→T 的颠换,4 个藏猪群体和八眉猪在第266 bp 处均为C,而长白猪和大白猪及烟台黑猪群体在该处存在C/T 杂合现象。说明分布在海拔较高(2 300 m以上)的地方猪种藏猪和八眉猪与海拔较低的烟台黑猪、长白猪和大白猪在EGLN1 基因上有着截然不同的遗传变异。
本研究在EGLN1 基因发现的变异位点界定的2 个等位基因(C 和T)及3 个基因型(CC、CT、TT)分布具有明显的海拔特征。在较高海拔的八眉猪和高海拔藏猪群体中只存在1 种等位基因C 和基因型CC。关于人的EGLN1 基因多态性研究也表明藏汉人群之间存在显著差异,这一结果为研究人类和动物在极端环境下的适应性进化提供参考[13]。在对藏族人群高原低氧适应性遗传机制研究中发现,EGLN1 基因有2 个SNP 位点,且因此界定的基因型和频率与汉族人差异极显著,并认为这种变异位点及频率之间差异的显著性与藏族人适应高原低氧环境存在相关性[14]。对藏绵羊和湖羊EGLN1基因多态性研究也表明,藏绵羊具有优势基因型和等位基因,且与湖羊基因频率差异显著,推测藏绵羊的优势基因型可能更有利于高原低氧适应[15]。而本研究中低海拔的大白猪、长白猪、烟台黑猪群与高海拔猪种之间等位基因、基因型频率及组成的特征性差异,推测可能与藏猪长期适应性进化中的环境选择有关。
