CYTOR通过调控miR-125b-5p/HK2通路促进宫颈癌细胞有氧糖酵解、增殖和侵袭
2019-08-12
(武汉市三医院妇产科,湖北 武汉430074)
随着手术、放疗、化疗等综合治疗方法的发展和改进,早期宫颈癌的疗效取得了很大的进展,但中晚期宫颈癌的疗效仍然不能令人满意[1]。中晚期宫颈癌的高度增殖活性和侵袭性直接导致了预后不良,因此,积极研究宫颈癌发生增殖和侵袭转移的机制,并寻求相应的治疗靶点,对于提高宫颈癌的整体疗效有十分明显的临床意义[1]。以往针对宫颈癌的靶向治疗主要集中在蛋白靶点,但近年来,一种新型的非编码RNA逐渐引起了肿瘤学者的重视。长链非编码RNA(long non-coding RNA, lncRNA)是一大类长度超过200个核苷酸的特殊RNA,由于不具备蛋白质编码能力长期未受到肿瘤研究者的重视。但近年来越来越多的研究表明,lncRNA在多种肿瘤中具有关键性的调节作用[2-3],跟蛋白靶点一样,lncRNA同样可以作为癌基因存在[4]。然而,关于lncRNA和宫颈癌之间的关系,以及lncRNA如何促进宫颈癌的增殖和侵袭活性,这方面的研究尚处于早期阶段。本研究首先通过检测一种新型lncRNA细胞骨架RNA(cytoskeleton regulator RNA, CYTOR)在宫颈癌组织及细胞中的表达,然后进一步探讨CYTOR通过调控miR-125b-5p和HK2对宫颈癌细胞增殖、侵袭和有氧糖酵解的影响,为寻求宫颈癌新的治疗靶点提供实验依据和理论支持。
1 材料与方法
1.1 组织、细胞株、试剂及仪器
选取来源于本院2014年5月至2018年5月的宫颈癌组织及癌旁组织30例,纳入标准:符合国际妇产科联合会(FIGO)分类标准且经病理确诊为宫颈癌,宫颈癌分期采用FIGO分期。4种人宫颈癌细胞株(Hela、C33A、Caski和SiHa)和人宫颈上皮细胞(H8)购于中科院上海细胞研究所。Si-CYTOR(sh1:CTGGAAACCTCTTGACTCT;sh2:CAGGAAGCTCTATGACACA)和pcDNA-HK2过表达载体(HK2全长序列见https://www.ncbi.nlm.nih.gov/nuccore/M23115.1)由上海吉玛制药技术有限公司合成,转染试剂Lipofectamine 2000购于Invitrogen公司;RNA提取试剂盒Trizol购于Invitrogen公司;实时定量SYBR Green和逆转录试剂盒购于TAKATA公司,引物由上海生工生物合成;HK2抗体和内参抗体购于Abcam公司;增殖检测CCK8试剂盒购于碧云天公司。双荧光素酶报告基因试剂盒购于Promega公司;Western blot用电泳仪和转印仪购于Bio-Rad公司。
1.2 细胞培养和转染
人宫颈癌细胞株(Hela、C33A、Caski和SiHa)和人宫颈上皮细胞(H8)培养于DMEM培养基中(培养基中含10%胎牛血清和青链霉素双抗),培养条件:温度37 ℃、含5%CO2。待细胞进入对数生长期,用胰酶消化后选取2 mL细胞悬液并接种到6孔板中(保持细胞密度为1×105个/mL),在培养箱培养24 h后按照Lipofectamine 2000说明书方法进行转染,转染物包括si-CYTOR、miR-125b-5p inhibitor和pcDNA-HK2,转染48 h后用于下一步检测实验。
1.3 荧光定量PCR检测CYTOR、miR-125b-5p和HK2的表达
收集宫颈癌组织及转染后48 h的宫颈癌细胞,采用TRIzol试剂提取宫颈癌组织和细胞总RNA,随后进行逆转录反应制备cDNA。荧光定量PCR检测方法以U6和GAPDH作为内参照,引物序列如下:U6 F为5′-CTCGCTTCGGCAGCACA-3′,R为5′-AAC GCTTCACGAATTTGCGT-3′;GAPDH F为5′-AATGGGCAGCCGTTAGGAAA3-3′,R为5′-GCGCCCAAT ACGACCAAATC′-3′;CYTOR F(2143-2162)为5′-AGAATGAAGGCTGAGGTGTG-3′,R为(2465-2484)5′-CAGCGACCATCCAGTCATTTA-3′;miR-125b-5p F为5′-TCCCTGAGACCCTAACTTGTGA-3′,R为5′-AG TCTCAGGGTCCGAGGTATTC-3′;HK2 F为5′-CAAA GTGACAGTGGGTGTGG-3′,R为5′-GCCAGGTCCTTCACTGTCTC-3′。按荧光定量试剂盒说明建立PCR反应体系(包括2 μL cDNA、10 μL SYBR Green Mix、上下游引物。PCR运行参数中设置:95 ℃ 3 min,然后进行下列反应:92 ℃变性30 s,60 ℃退火30 s,一共进行35个循环。PCR检测结果采用2-ΔΔCt法进行汇总分析。
1.4 宫颈癌细胞的增殖活性的检测(CCK-8法)
培养HeLa细胞使之处于对数生长期,然后接种细胞于96孔板中(保持细胞密度为2×104个/孔),细胞中加入100 μL DMEM培养基,每组设立3个平行对照。在HeLa细胞中分别转染si-CYTOR、pcDNA-HK2和miR-125b-5p inhibitor。检测前加入10 μL CCK-8溶液,然后将细胞在培养箱内孵育4 h后测定450 nm处的光密度OD值。所有操作依照CCK-8试剂盒说明书进行。
1.5 Transwell法检测宫颈癌细胞的侵袭能力
将HeLa细胞用胰酶消化后接种于Transwell小室内,Transwell上室加100 μL HeLa细胞悬液,下室加250 μL培养基(含10%胎牛血清),在37 ℃、5%CO2环境培养箱中培养48 h,取出小室后用PBS溶液冲洗2遍,然后用4%多聚甲醛固定15 min,最后用结晶紫溶液进行染色,Transwell板干燥后置于倒置显微镜下观察细胞侵袭情况。
1.6 Annexin V-FITC/PI检测细胞凋亡
选取HeLa细胞并培养至对数生长期,用PBS清洗后均匀地混合HeLa细胞与500 μL预冷的结合缓冲液和5 μL Annexin-V-FITC,避光放置15 min,上机前5 min加入2.5 μL PI溶液进行染色,均匀混合后上流式细胞仪检测细胞凋亡率。所有操作依照Annexin V-FITC/PI试剂说明书进行。
1.7 双荧光素酶报告基因验证miR-125b-5p和CYTOR/HK2之间的互作关系
首先将CYTOR 3′UTR靶序列或HK2 3′UTR靶序列插入到荧光素酶载体下游。并设置突变载体作为对照进行荧光素酶验证实验,具体操作如下:每孔细胞中转染pmir-GLO-CYTOR-3′UTR 200 ng、pmir-GLO-HK2-3′UTRmiRNA、mimic 500 ng及阴性对载体200 ng。用Opti-MEM培养基将Lipofectamine2000稀释后,室温下放置5 min;然后混合转染试剂和载体DNA和mimic,室温下放置20 min后,用Opti-MEM培养基进行充分混匀;然后在温度37 ℃、5%CO2培养箱中培养48 h后收集细胞。整个检测步骤依照双荧光素酶报告基因试剂盒说明书,并以载体本身的荧光值作为内对照。
1.8 统计学分析
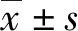
2 结 果
2.1 CYTOR在宫颈癌组织及细胞中表达升高
首先,利用荧光定量PCR探明CYTOR在宫颈癌组织和细胞中的表达,结果显示,相比癌旁组织(图1A),CYTOR在宫颈癌组织中的表达明显增高(3.29±2.21vs8.39±4.86,t=6.83,P<0.01);对30例宫颈癌患者的CYTOR和临床病理特性进行了分析,结果见表1所示,分期是导致CYTOR表达差异的重要因素。荧光定量PCR结果表明,相比I/II级宫颈癌组织(图1B),CYTOR在III/IV宫颈癌中的表达也明显升高(5.25±2.02vs9.77±5.12,t=3.99,P<0.01)。同时细胞实验表明,相比人正常宫颈上皮细胞H8,CYTOR在宫颈癌细胞系(Hela、C33A、Caski和SiHa)中的表达水平明显增高(t=19.71、16.23、-19.38、20.39,P<0.01,图1C)。以上结果显示,CYTOR的异常表达升高可能与宫颈癌的恶性生物学行为存在密切关联。
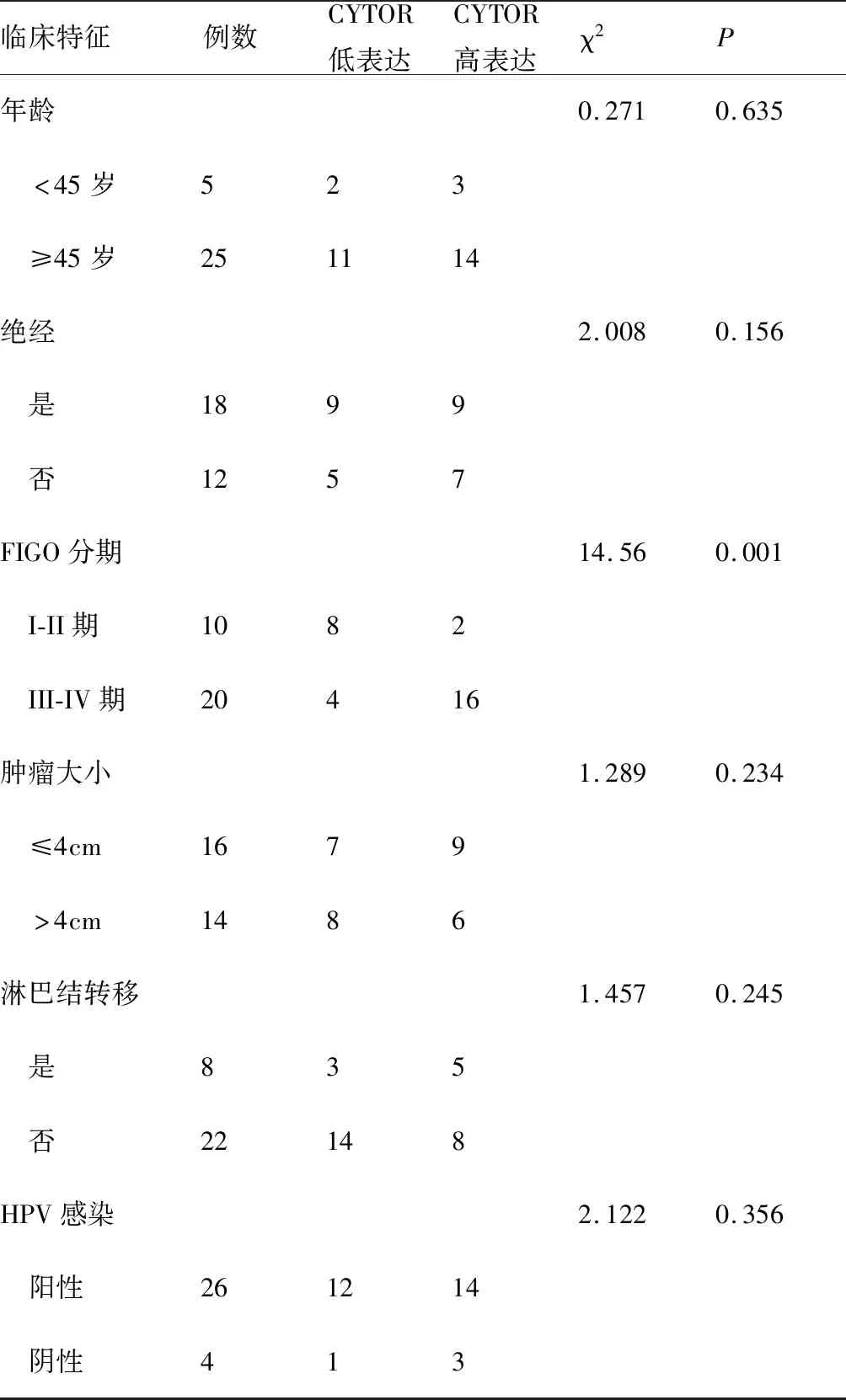
表1 30例宫颈癌患者临床病例特征与CYTOR表达的关系
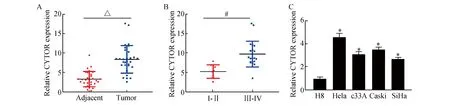
图1 CYTOR在宫颈癌组织和细胞中的表达A:CYTOR在宫颈癌组织和癌旁组织中的表达;B:CYTOR在I/II级宫颈癌以及III/IV宫颈癌中的表达;C:CYTOR在宫颈癌细胞以及正常上皮细胞中的表达 与Adjacent比较,△P<0.05;与Ⅰ/Ⅱ比较,#P<0.05;与H8组比较,*P<0.05
2.2 抑制CYTOR对宫颈癌细胞增殖和侵袭的影响
荧光定量PCR法检测结果(图2A)显示,相比于对照组,转染CYT OR siRNA可明显下调CYTOR在宫颈癌细胞HeLa中的表达(P<0.05),且转染si-CYTOR#1的效果优于si-CYTOR#2和si-CYTOR#3,在接下来的实验中选用si-CYTOR#1。CCK-8检测结果显示(图2B),抑制CYTOR可抑制HeLa细胞的增殖活力(t=7.23,P<0.01)。Transwell检测结果显示(图2C),抑制CYTOR可抑制HeLa细胞的侵袭能力(t=8.85,P<0.01)。此外,细胞凋亡结果显示(图2D),抑制CYTOR可促进HeLa细胞凋亡(t=12.78,P<0.01)。以上实验结果显示,抑制CYTOR可抑制HeLa细胞的增殖和侵袭能力并促进其凋亡。
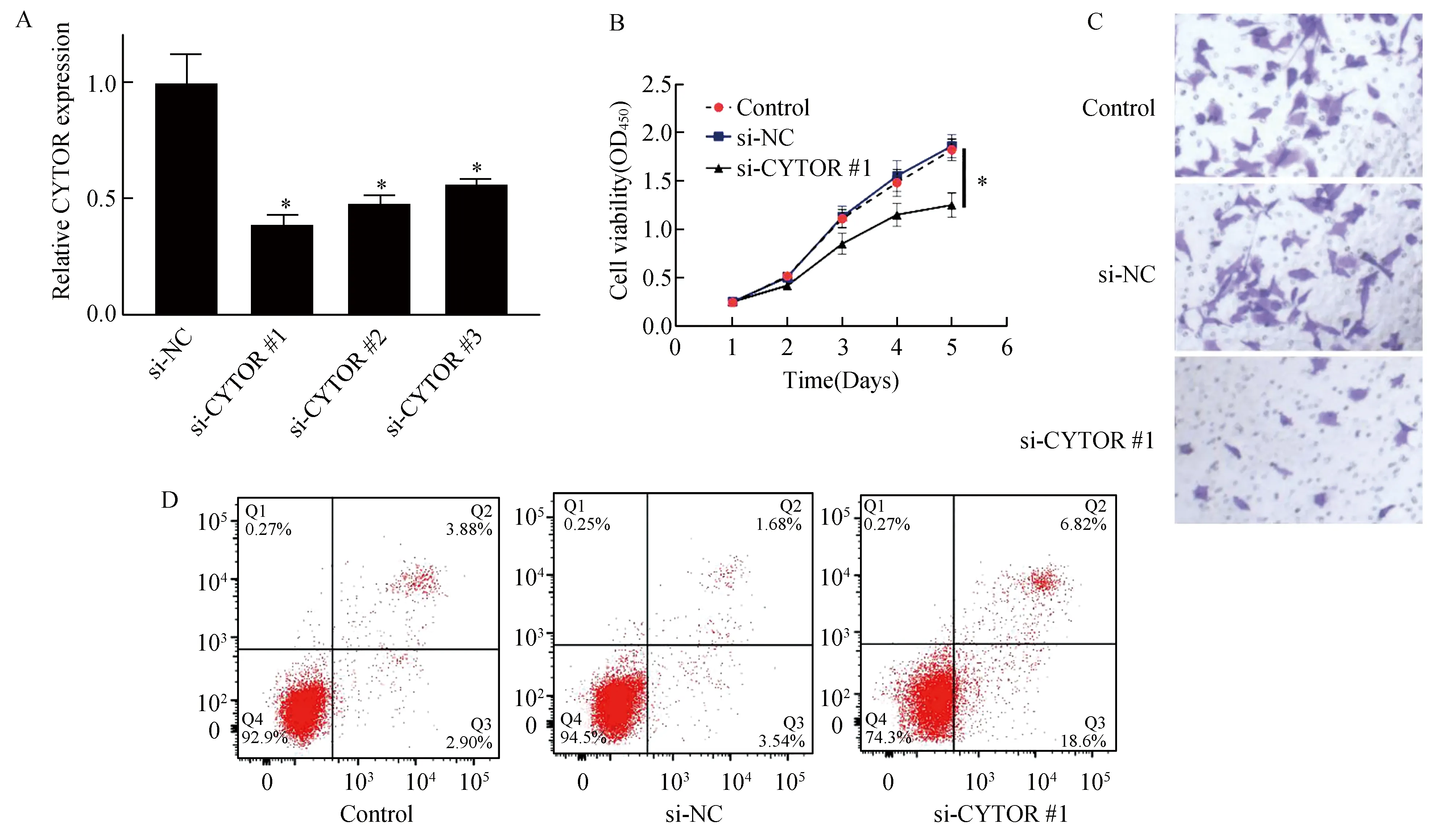
图2 抑制CYTOR后对宫颈癌细胞增殖、侵袭及凋亡的影响A:CYTOR表达量的变化;B:对宫颈癌细胞增殖;C:对宫颈癌细胞侵袭;D:宫颈癌细胞凋亡与si-NC组比较,*P<0.05
2.3 抑制CYTOR对宫颈癌细胞有氧糖酵解代谢的影响
肿瘤的有氧糖酵解能力增强是肿瘤获得快速增殖和侵袭的重要机制,为了探明CYTOR对宫颈癌细胞生物学功能影响的具体机制,对有氧糖酵解活性进行了检测,结果表明,抑制CYTOR后,无论是对葡糖糖的摄取水平(图3A),还是乳酸(图3B)和三磷酸腺苷(adenosine triphosphate,ATP)(图3C)的生成水平均发生了明显下降(t分别为6.86,7.52,6.95,P<0.01),表明CYTOR对维持宫颈癌细胞的有氧糖酵解水平有重要作用。
2.4 CYTOR靶向调控miR-125b-5p在宫颈癌细胞中的表达
利用生物信息学数据库Stabase V3.0数据库(http://starbase.sysu.edu.cn/)预测miR-125b-5p可能是CYTOR的靶基因,其预测序列如图4A所示。双荧光素酶报告基因验证结果(图4B)显示,过表达miR-125b-5p可使荧光素酶活性显著下降(t=6.58,P<0.05),而共转染miR-125b-5p mimics和靶向位点发生突变的pmirGLO-CYTOR-mut载体,miR-125b-5p对荧光素酶活性未发生明显作用。采用荧光定量PCR检测CYTOR抑制后miR-125b-5p在Hela中表达水平,结果(图4C)显示,CYTOR抑制后显著促进miR-125b-5p的表达(t=16.52,P<0.01)。通过对30例宫颈癌标本进行检测发现,CYTOR与miR-125b-5p的表达呈负相关(r2=0.377,P<0.01,图3D)。由此可知,miR-125b-5p与CYTOR存在直接作用,CYTOR可负调控miR-125b-5p的表达。
2.5 miR-125b-5p靶向调控HK2在宫颈癌细胞中的表达
通过生物信息学数据库Stabase V3.0对miR-125b-5p的靶基因进行了预测,发现HK2是miR-125b-5p的候选靶基因,随后采用荧光素酶报告基因验证,miR-125b-5p可以结合HK2的3′UTR靶序列(图5A),并且miR-125b-5p可以抑制HK2的表达(t=7.32,P<0.05,图5B)。采用荧光定量PCR检测miR-125b-5p抑制后对HK2 mRNA表达的影响,结果显示,miR-125b-5p抑制后可显著促进HK2 mRNA在Hela细胞中的表达(t=18.35,P<0.01,图5C)。同样对30例宫颈癌组织标本进行检测,结果显示miR-125b-5p与HK2的表达呈负相关(r2=0.265,P<0.01,图5D)。上述结果表明,HK2是miR-125b-5p的靶基因,同时miR-125b-5p可负调控HK2的表达。
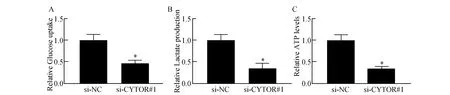
图3 抑制CYTOR后对宫颈癌细胞有氧糖酵解能力的影响A:宫颈癌细胞葡糖糖摄取;B:宫颈癌细胞乳酸生成;C:宫颈癌细胞ATP生成与si-NC组比较,*P<0.05
3 讨 论
通常认为,包括宫颈癌在内的恶性肿瘤的发生,内在癌基因的促进起到了十分关键的作用。因此,在过去几十年,对癌基因及以其为靶点的研究成为热门,传统的观点认为,癌基因就是蛋白质,因此恶性肿瘤的发生与蛋白质靶点的突变和异常相关,这使得研究者的焦点主要集中在基因编码即编码RNA领域,而忽视了占转录本上绝大部分的非编码RNA,使得非编码RNA产物在以往通常被称之为“转录暗物质”。但近年来,随着对基因表达调控尤其是表观遗传学的深入研究,非编码RNA也是影响恶性肿瘤发生和发展的重要因素甚至是决定性因素。目前,非编码RNA主要分为小分子RNA(microRNA)、环状RNA(circular RNA, circRNA)、转运RNA(transfer RNA,tRNA)和长链非编码RNA(long non-coding RNA,LncRNA)等几大类别,其中尤以LncRNA受到特别的关注。LncRNA是一大类长度超过200 bp的转录本,作为一种近年来新发现的非编码RNA,被认为在调控恶性肿瘤的生物学行为中起到了关键作用,例如LncRNA参与了恶性肿瘤的发生、增殖、侵袭、转移、抗凋亡、耐药等诸多恶性表型[4]。例如在宫颈癌中,lncRNA HOXD簇反义RNA 1(HOXD cluster antisense RNA 1,HOXD-AS1)可以促进宫颈癌细胞的增殖[5];同样的,结直肠肿瘤差异表达基因(Colorectal Neoplasia Differentially Expressed,CRNDE)也被认为是一个促进宫颈癌增殖和侵袭活性的癌基因[6]。CYTOR是一个近年来新发现的一个lncRNA,被认为在多种恶性肿瘤中发挥作用,例如在非小细胞肺癌中,CYTOR可以促进肿瘤细胞的增殖、侵袭并促进对放射治疗的抗拒性[7]。又比如在结直肠癌细胞中,CYTOR可以促进肿瘤细胞的增殖,被认为是一个癌基因[8]。但目前为止,CYTOR在宫颈癌中的作用及机制如何,尚未见报道。本研究发现,CYTOR在宫颈癌组织和细胞中高表达,揭示了CYTOR在促进宫颈癌恶性表型中的作用。同时功能实验证实,抑制CYTOR的表达可以明显抑制宫颈癌细胞的增殖和侵袭活性,同时宫颈癌细胞的有氧糖酵解能力也显著下降。这些实验表明,CYTOR很可能是通过调节有氧糖酵解能力来调控宫颈癌细胞的增殖和侵袭活性的。
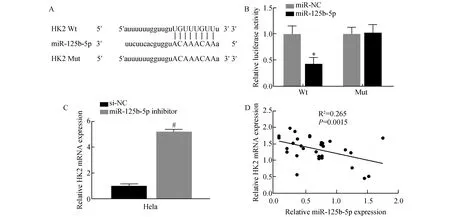
图5 miR-125b-5p靶向调控HK2的表达A:HK2 mRNA 3UTR与miR-125b-5p存在互补关系;B:荧光素酶实验证实HK2 mRNA与miR-125b-5p存在相互作用;C:抑制miR-125b-5p对HK2表达的影响;D:直线相关分析证实HK2与miR-125b-5p存在线性表达关系与miR-NC组比较,*P<0.05,与si-NC比较,#P<0.05
LncRNA可以通过多种途径调控基因功能,例如与靶基因的启动子区域结合调控靶基因转录,也可直接与信使RNA(messenger RNA,mRNA)结合调控靶基因的稳定性,但目前研究LncRNA的焦点还是集中在“分子海绵”这一独特作用上。LncRNA可以特异性地与相应microRNA相结合,从而发挥“分子海绵”的作用影响microRNA对靶基因的调控[9]。例如UCA1通过吸附miR-206从而促进宫颈癌的转移活性[10]。在本研究中,通过生物信息学和荧光素酶互作实验证实,CYTOR可以竞争性吸附miR-125b-5p来促进癌基因HK2转录。miR-125b-5p被认为是一个抑癌基因,在多种恶性肿瘤中发挥抑癌作用。例如,miR-125b-5p可以抑制乳腺癌细胞的增殖和侵袭转移[11];同样的,在喉癌细胞中,miR-125b-5p可以抑制肿瘤细胞的增殖,其高表达可以提示喉癌患者具有良好的预后[12]。本研究也首次证实了miR-125b-5p在宫颈癌细胞中同样发挥抑癌基因的作用,抑制CYTOR导致的miR-125b-5p活性提高可以抑制宫颈癌细胞的增殖和侵袭能力。因此,宫颈癌细胞中CYTOR的高表达,可能通过竞争性下调miR-125b-5p的活性,从而发挥癌基因的作用。
维持能量供给是细胞存活的先决条件之一,不同于正常细胞主要通过线粒体氧化途径来供能,包括宫颈癌细胞在内的多种癌细胞,即使在氧气充足的条件下,依然优先通过有氧糖酵解途径来供能,这种特殊的有氧糖酵解活性,被认为是恶性肿瘤的十大重要特征之一[13]。因此,靶向有氧糖酵解通路,或者靶向调控有氧糖酵解的关键酶,被认为是抗肿瘤研究的热门方向。HK2作为影响糖酵解途径的关键限速酶之一,参与了多种肿瘤的发生发展,是一个潜在的肿瘤治疗靶点[14]。本研究通过荧光素酶实验证实,HK2可以与CYTOR/miR-125b-5p轴发生直接相互作用,CYTOR可能促进HK2的表达来促进宫颈癌细胞的增殖和侵袭活性,同时也增加了宫颈癌细胞的有氧糖酵解能力。同时表明,CYTOR/miR-125b-5p/HK2这一全新分子轴在宫颈癌的增殖侵袭以及有氧糖酵解调控上发挥重要作用。
综上所述,本研究通过功能实验证实,lncRNA CYTOR通过下调miR-125b-5p来上调HK2表达进而促进宫颈癌细胞增殖、侵袭和有氧糖酵解活性,从而促进宫颈癌的恶性表型。研究结果揭示CYTOR/miR-125b-5p/HK2有可能成为宫颈癌治疗的有效靶点。但更具体的分子机制和靶向宫颈癌治疗效果,有待今后进一步更细致深入的分子通路和动物实验来证实。
