珠子参表型性状变异及其与地理-土壤养分因子的相关性分析
2019-08-08左应梅杨维泽杨美权杨绍兵杨天梅许宗亮李纪潮张金渝
左应梅,杨维泽,杨美权,杨绍兵,杨天梅,许宗亮,李纪潮,张金渝
(云南省农业科学院药用植物研究所,云南 昆明 650205)
【研究意义】珠子参,又名扣子七、纽子七、珠儿参,为五加科人参属植物珠子参(PanaxjaponicusC.A. Mey. var. major C. Y. Wu et K. M. Feng)或羽叶三七[PanaxjaponicasC. A. Mey. var.bipinnatifidus(Seem.) C. Y. Wu et K. M. Feng]的干燥根茎[1]。珠子参气微,味苦微甘,主治气阴两虚,虚劳咳嗽,跌扑损伤等[2],且在心血管、中枢神经和免疫系统等方面也有不同程度的药理活性[3]。由于珠子参具有较高的药用价值,促其商品价格不断攀升,而市场的高价位吸引人们无序采挖,导致珠子参产区植被生态系统严重破坏,其种群资源出现濒危化趋势[1]。因此,有必要针对珠子参野生资源的保护和合理开发利用开展相关研究。【前人研究进展】植物的表型是由遗传因素和环境因素共同决定的[4],是基因和所处生态环境共同作用的结果,是生物遗传变异的表征。而天然居群的表型变异具有适应和进化上的意义,是人工驯化和遗传育种研究的基础,也是揭示种群适应性的有效途径之一。近年来,国内外学者对鹅观草[5]、扇脉杓兰[6]、暴马丁香[7]]等多种植物的表型变异与环境因子间的相关性进行了研究报道。目前,珠子参的研究主要集中在资源分布[1]、生物学特性[8]、栽培管理[9]、化学成分[10]、药理活性[11]、遗传多样性[12]和分子生物信息学[13]等方面。【本研究切入点】但关于其表型与生理特征的地理变异的研究较少。【拟解决的关键问题】以云南省内的12个珠子参天然居群为对象,研究珠子参的株高、茎粗、叶长/宽等表型性状与环境因子间的关系,以期了解珠子参种源的遗传变异程度、揭示其变异规律,为珠子参种质资源的收集和保存、优良种源的选择和遗传改良以及良种选育提供理论依据。
1 材料与方法
1.1 性状测定方法和样本采集
2015年7月20-26日,分别对12个珠子参天然居群进行实地调查。详细观察和记录珠子参居群内的表型特征以及群落所处的自然条件、群落特点及生境,其中经度、纬度及海拔采用GPS测定,光照强度采用SAUTER SO 200 K便携式光照强度测量仪(德国)测定。每个居群选取20个植株,测量株高(无花株高的测量为植株基部地面处到最高叶面的距离,有花植株株高的测量为地表到植株最高处的距离)和茎粗;在每一植株中部选取完整的叶片 3 片,分别测定叶长/宽、小叶柄长和复叶柄长,其中,叶长为叶尖与小叶柄基部之间的长度,叶宽为叶片中部的宽度,复叶柄长为小叶柄基部与茎杆之间的长度。以上指标,茎粗用游标卡尺测量,其余指标均用钢卷尺测量。所有样品经云南省农业科学院药用植物研究所张金渝研究员鉴定为野生珠子参。
1.2 土壤因子测定
在每个样地内确定 3 个样点,挖取距地表深 0~20 cm 根系附近土壤,将3个样点的土壤样品混合,自然风干后研磨、备用。用于检测pH值、有机质、氮、钾、磷以及全量铜、锌、铅、铬、镉、镍、砷、汞等指标。其中pH值:电极法;有机质:重铬酸钾容量法;有效磷:碳酸氢钠浸提-钼锑抗比色法;速效钾:中性醋酸铵浸提-火焰光度法;全氮含量:半微量开氏法;磷含量:钼锑抗比色法;全钾含量:火焰光度计法。土壤重金属 Pb、Cr、Cu 和 Zn 含量:使火焰原子吸收分光光度计测定;Cd 含量,使用石墨炉原子吸收分光光度计测定;As 和Hg 含量:使用原子荧光分光光度计测定[14]。
1.3 数据统计分析
采用Excel 2007进行数据的处理,采用JMP7.0软件对数据进行方差分析、多重比较和相关分析;采用SPSS 22.0对12个珠子参天然居群的7个表型性状进行系统聚类分析(组间联接法)。
2 结果与分析
2.1 珠子参12个天然居群的分布情况
调查点分布于云南省滇西北,北纬26°56′39.65″~28°27′46.41″,东经98°55′37.19″~100°11′30.66″,海拔范围在2795~3597 m。生态环境以阔叶灌木、竹叶林下居多,也有生长在针阔叶林下,遮阴度在0~90 %,早晨至中午有一定散射光的西北向和东北向山坡、河谷坡地生长较好,伴生植物多为蕨类、竹类、升麻等(表1)。土壤主要为黑褐色腐殖质土,土壤的有机质、氮、钾等含量丰富,有效磷含量相对偏低(表2)。土壤中的重金属含量基本未超过《土壤环境质量标准》(GBl5618-1995) 中的二级标准,表明野生珠子参分布地的土壤质量状况良好,重金属含量相对较低(表3)。
2.2 珠子参12个天然居群的变异特征
由表4可知,珠子参12个天然居群的数量性状茎数在居群间与居群内没有变异,而小叶数在居群间与居群内存在明显变异;茎色、叶形、叶缘等质量性状在居群间与居群内均存在明显变异。表型性状在居群间与居群内也存在广泛变异,其中茎粗和复叶柄长在居群间的变异最大,变异系数的平均值分别为69.18 %和57.49 %;12个居群表型性状的平均变异系数最大的5个居群依次为:香格里拉普达措(PDC1)53.61 %>香格里拉普达措(PDC2)52.04 %>丽江文海(WH)42.9 %>德钦隧道(DQSD)42.56 %>玉龙县拉美荣(LMR2)40.81 %,平均变异系数最小的居群是虎跳峡尾木尼村(WMN)27.11 %(表5)。比较12个天然居群表型性状(平均值),以茎粗和复叶柄长在居群间差异最大(表6)。

表1 珠子参12个天然居群的生境条件

表2 珠子参12个天然居群样地的土壤养分含量

表3 珠子参12个天然居群样地的土壤重金属含量
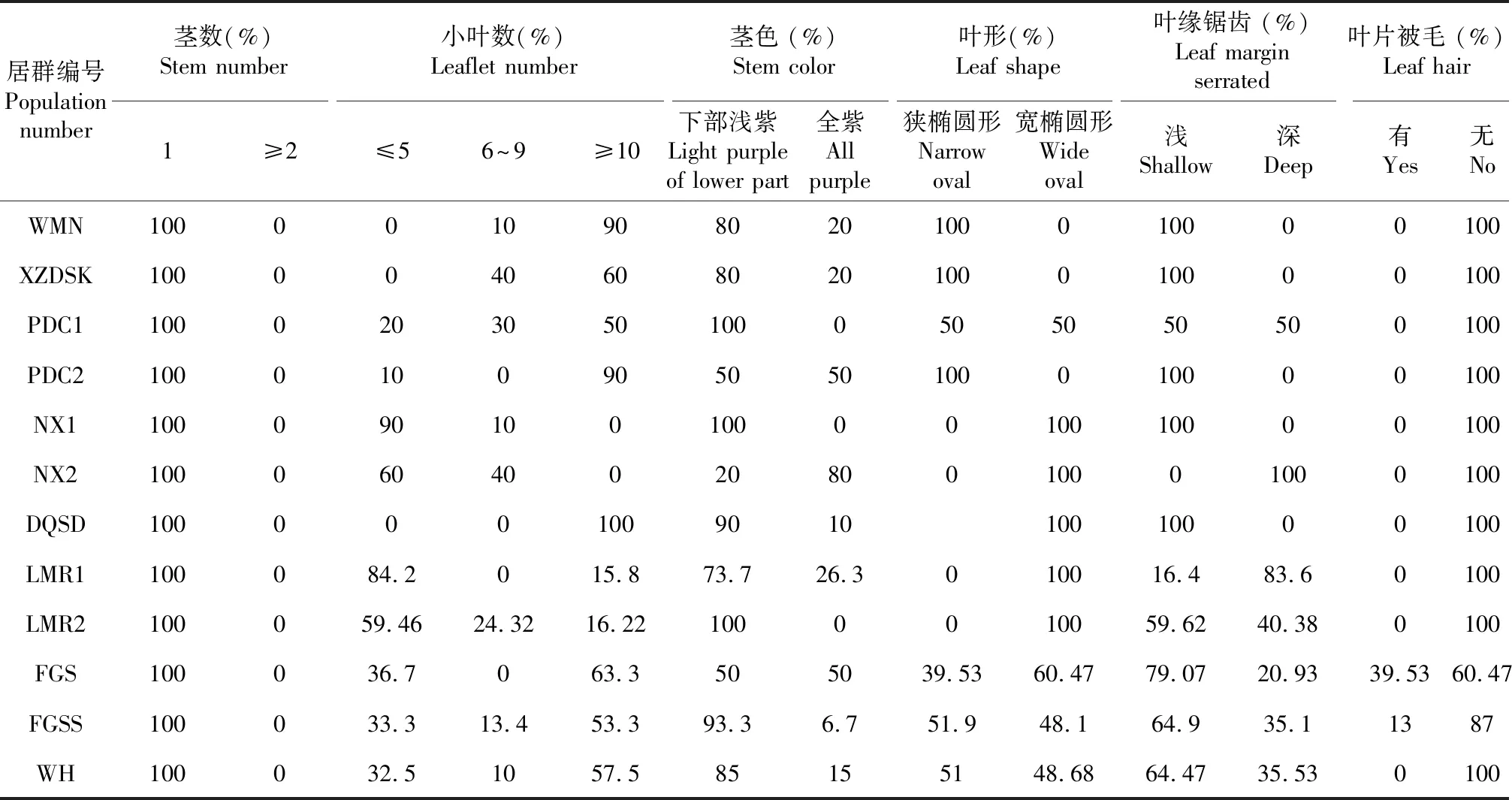
表4 珠子参12个天然居群的数量及质量性状频度
2.3 珠子参表型与地理-土壤养分因子间的关系
相关性分析表明,珠子参居群的叶长、复叶柄长与纬度和海拔存在显著负相关关系;小叶柄长与经度存在显著正相关关系;株高、叶长、小叶柄长与土壤中的有效磷含量存在显著正相关关系(表7)。
2.4 珠子参各居群间的相似性分析
为了研究珠子参居群间的相似性及亲缘关系,根据其形态特征的7个表型性状对12个珠子参天然居群用SPSS软件进行聚类分析。结果显示,12个天然居群在阈值为5时可明显分为4类,其中居群NX1、NX2和LMR1聚为一类;LMR2和FGSS聚为一类;WMN、DQSD、 XZDSK、FGS、PDC1和 PDC2聚为一类;WH单独聚为一类。由此可见,性状的表型特征并没有完全依地理距离而聚类,说明珠子参居群间表型性状变异存在不连续性。

表5 珠子参12个天然居群表型性状的变异系数
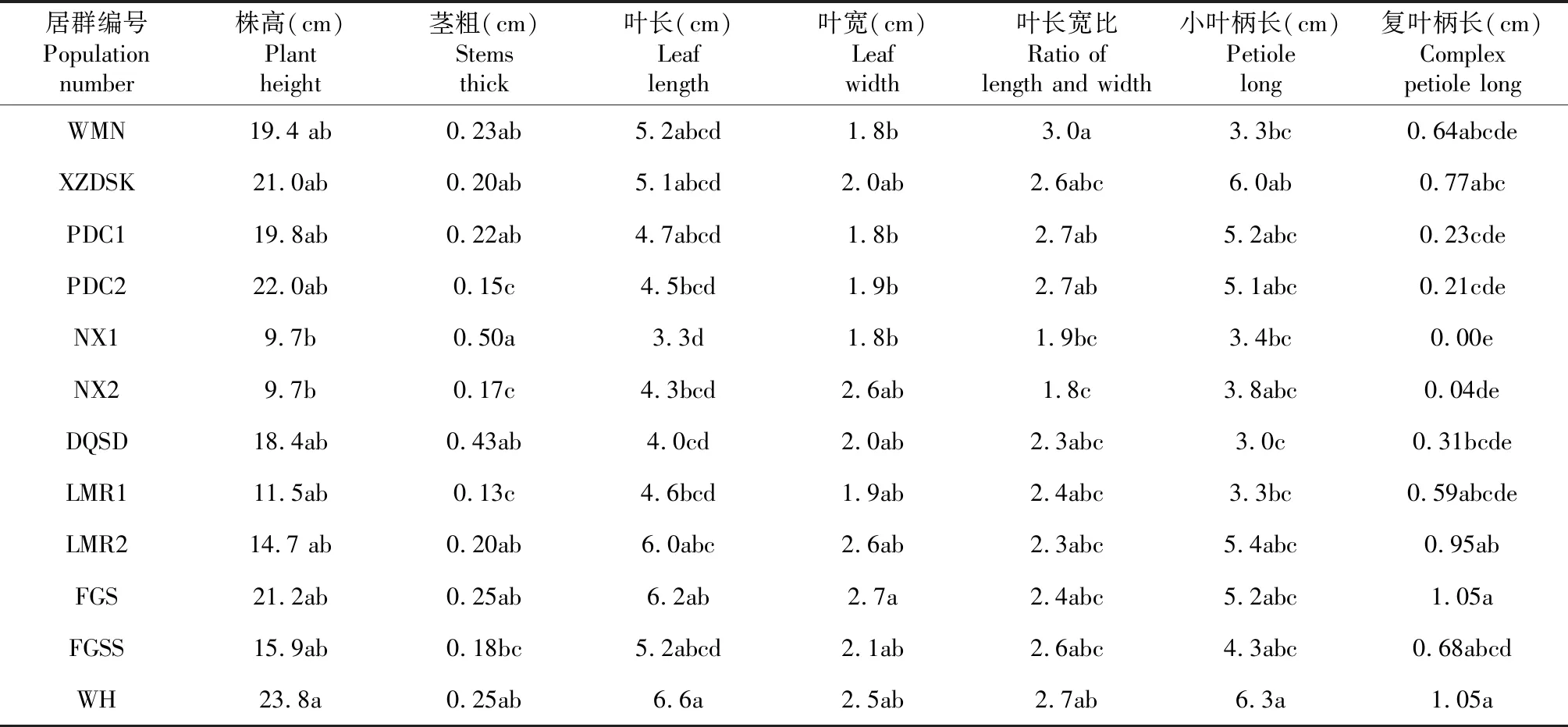
表6 珠子参12个天然居群表型性状的比较
注:同列数据后不同小写字母表示0.05水平差异显著。
Note: The data in the same column shows significant difference at 5 % level.
3 讨 论
植物表型性状的变异在适应性和进化上均具有非常重要的意义[15]。种源间变异能够反映植物在地理生殖隔离上的变异,是种内多样性的重要组成部分[16]。植物种群内遗传多样性或变异越丰富,则越有利于种群适应环境和进化。本研究对珠子参12个天然居群植株的数量性状、质量性状及表型性状的调查研究发现,数量性状小叶数在居群间与居群内存在明显变异,茎色、叶形、叶缘等质量性状在居群间与居群内也存在明显变异,这与杨天梅等[17]研究的珠子参的叶形态在同一居群内存在显著变异的研究结果相符。居群间各表型性状变异系数的平均值为20.28 %~69.18 %,表型变异非常丰富,其中茎粗和复叶柄长在居群间的变异最大,变异系数的平均值分别为69.18 %和57.49 %;对12个天然居群表型性状的平均值进行比较,茎粗和复叶柄长在居群间差异最大,这表明,珠子参的表型性状茎粗和复叶柄长对环境因子变化的敏感性高,在居群间的稳定性低。由上可知,珠子参的表型性状在居群间和居群内均存在着丰富的变异,说明珠子参有较强的环境适应性,这与段青等[18]在泸定百合上的研究结果相似。但是对于珠子参表型变异是否受基因控制以及受控程度尚不清楚,有待进一步研究。

表7 各表型性状与地理-土壤养分因子的相关性
注:*表示在 0.05 水平上差异显著。
Note: * means significant difference at 0.05 level.

图1 珠子参12个天然居群聚类图Fig.1 The cluster map of 12 natural populations of P. japonicus
有研究表明,植物的表型性状与地理-土壤养分因子存在显著的相关性,如王丹等[19]研究表明,茶条槭的叶长/宽、叶柄长等17个表型性状与土壤中的全氮、全钾、速效氮、速效钾、有效磷、土壤有机质及土壤含水量都存在显著或极显著的正、负相关性。肖海峻等[5]研究表明,鹅观草的株高、穗长、旗叶长/宽、小穗数、小花数穗下第1节间长和穗轴第1节长等8个性状与海拔、纬度和年平均温的相关性较高。李全健等[6]研究表明,扇脉杓兰野生居群表型性状存在丰富的变异和多样性。扇脉杓兰的地径与速效钾含量呈显著正相关,子房长、唇瓣长和宽均与土壤中全磷的含量呈显著正相关,植株展度与土壤 pH 呈显著正相关。杨晓霞等[7]研究表明,不同产地暴马丁香各表型性状间及表型性状与地理-气候因子间均存在不同的相关性,其中,种子发芽率与种子的其他5 个表型性状间的相关性均不显著;经度和纬度与各表型性状间的相关性不显著,而无霜期、年平均气温和海拔则是影响表型性状变异的主要因子。本研究中,珠子参居群的叶长、复叶柄长与纬度和海拔存在显著负相关关系;小叶柄长与经度存在显著正相关关系;株高、叶长、小叶柄长与土壤中的有效磷含量存在显著正相关关系,说明土壤中有效磷的含量是导致珠子参居群表型性状多样性的原因之一。以上分析表明,物种之间表型性状与地理因子及土壤养分因子之间的相关性存在差异,可能与其生境中的土壤肥力或各居群生境的异质性有关。
聚类分析结果表明,12个珠子参天然居群可初步分为4类,但性状的表型特征并没有完全依地理距离而聚类。这说明珠子参居群间表型性状变异存在不连续性,与宋杰等[20]研究得出的云南含笑6个天然居群的表型性状并没有严格依地理距离而聚类的结果类似。
4 结 论
珠子参的小叶数、茎色、叶形、叶缘等性状在居群间与居群内均有明显变异;珠子参居群的叶长、复叶柄长与纬度和海拔存在显著负相关关系;小叶柄长与经度存在显著正相关关系;株高、叶长、小叶柄长与土壤中的有效磷含量存在显著正相关关系,说明土壤中有效磷的含量是导致珠子参居群表型性状多样性的原因之一。12个珠子参天然居群可归分为4类,但性状的表型特征并没有完全依地理距离而聚类,说明珠子参居群间表型性状变异存在不连续性。
