辛伐他汀对BMSCs成骨分化过程中WNT信号通路早期WNT3的影响
2019-08-07贾忠宝崔纳尹丹丹田发明张柳
贾忠宝 崔纳 尹丹丹 田发明 张柳
1.北京市丰台区南苑医院骨外科,北京 100076 2.北京市丰台区南苑医院功能科,北京 100076 3.山东省潍坊市人民医院病理科,山东 潍坊 261599 4.华北理工大学附属医院医学实验中心,河北 唐山 063000 5.华北理工大学附属医院骨外科,河北 唐山 063000
多项研究证实,骨髓基质干细胞(bone marrow stromal cells,BMSCs)在不同诱导条件下具有多向分化潜能比如成骨细胞等分化[1]。多种因素及因子调控了BMSCs的分化潜能,在这其中不同特异性标志物表达水平也有不同的改变,如因子WNT3、β-catenin及Runx2、ALP、Ⅰ型胶原等。经典而又十分保守的Wnt信号传导通路的主要有配体(Wnt家族分子)、跨膜受体(Frizzled家族分子和LRP-5/6)、胞浆调节蛋白(Dsh、β-catenin、APC、Axin、GSK-3β等)以及核内转录因子(TCF/LEF1家族)等组成。
Wnt信号通路增加骨量是通过不同的机制来实现,研究发现在调控骨形成发育、生长及骨量增加过程中,经典的Wnt/β-catenin信号通路以时空形式发挥着重要作用[2-3]。BMSCs向成骨细胞、软骨细胞及脂肪细胞的不同分化结果主要是因为在细胞分化的不同阶段、时期,Wnt/β-catenin信号通路在多种调剂机制下发挥了不同的作用。
研究发现,具有多条密切相关的信号传导通路参与到BMSCs向成骨细胞复杂的分化过程中,既往研究多侧重于转化生长因子信号传导途径[4],新的研究证实在成骨细胞的分化及骨折的愈合过程中Wnt经典信号传导途径至关重要[5-6]。而Wnt蛋白将信号传入细胞内必须要与frizzled受体结合。这条Wnt/β-catenin途径称为“经典途径”[7]。Wnt信号通路激活时, GSK-3β被募集并磷酸化LRP6,而不能磷酸化β-catenin从而避免被其降解;但在Wnt信号未激活时,GSK-3β则磷酸化β-catenin,并促使其降解。
辛伐他汀在SD大鼠骨髓基质干细胞在WNT信号通路调节下向成骨细胞分化过程中早期的作用如何,相关的研究文章国内外较少报道发表。辛伐他汀早期干预可以促进骨髓基质干细胞成骨细胞分化过程中骨钙素水平的增高[8]。可本研究采用在第二代细胞中给药的方法在早期分别提取RNA和蛋白来分析骨髓基质干细胞定向诱导分化为成骨细胞培养过程中辛伐他丁所发挥的作用及对Wnt信号通路的影响,以进一步探讨促进骨形成的机制。
1 材料和方法
1.1 材料
1.1.1动物:4周龄雌性SD大鼠10只(北京大学提供SPF级,合格证号SCXK(京)2006-0 008号)。10只SD大鼠均安实验要求单笼适应环境饲养1周。
1.1.2试剂和仪器:由Sigma 公司购买DMEM培养基、辛伐他汀、维生素C、β-甘油磷酸钠、氢化可的松及血清。由大连宝生物公司购买Realtime PCR 试剂盒及Trizol (Invitrogen),引物由Invitrogen公司合成。
1.2 方法
1.2.1提取及体外培养骨髓基质干细胞:取4周龄SD大鼠的股骨和胫骨。收集由培养基冲洗出来的骨髓细胞于离心管中,以1 000 r/min的速度离心10 min,弃上清,应用完全DMEM培养液重悬吹打后每只大鼠的细胞接种于一个培养瓶中,拧松瓶口并在37 ℃恒温箱中以含有50 mL/L(体积分数)的二氧化碳环境下培养2 d后换液,后期视情况每间隔2~3 d更换一次培养液。待细胞贴壁并增殖至培养瓶底的4/5后加入诱导培养基,待细胞增殖融汇成致密单层后再传代接种于新的培养瓶。
1.2.2分组与加药:在第二代细胞第一次换液时取生长良好的细胞更换诱导培养基并随机分组,分为对照组(G1)和实验组(G2)。G2加入诱导培养基(含有1×10-7mol/L SIM),对照组加入含有等量DMSO的培养基。以后视细胞生长情况间隔2~3 d全量换液,每次换液均要求实验组加入含有辛伐他丁的诱导培养基,G1加空白完全培养,分别在12 h和36 h,提取总RNA和蛋白质。
1.2.3Real-time RT-PCR:选取第二代细胞在经诱导分化12 h和36 h后两组分别提取细胞总RNA,定量后经过反转录合成第一链cDNA进行PCR反应,同批扩增待测基因与内参基因(β-actin),采用溶解曲线法进而来鉴定扩增产物的特异性和片段大小,应用梯度稀释内参基因法制定相对标准曲线。PCR反应体系的总体积为25 μL,其中上下游引物各0.5 μL、12.5 μL的STBR® Premix Ex TapTM,2 μL的DNA模板及dH2O 9.5 μL。PCR反应条件:在经过95 ℃ 30 s、95 ℃ 5 s、60 ℃ 30 s,共计40个循环。对RT-PCR结果进行半定量分析。查出WNT3的cDNA全长序列后应用软件Primer 5.0设计因子的引物并由Invitrogen公司合成。详见表1。

表1 WNT3及β-actin引物序列Table 1 List of PCR primers
1.2.4Weten blot:选取第二代骨髓基质干细胞在经诱导培养后用辛伐他汀干扰12 h和36 h,两组细胞分别提取总蛋白,以50 μg/孔的标准上样行凝胶电泳进行蛋白分离;分离后将蛋白转移至PVDF膜于BSA中在室温环境下封闭2 h,将膜与一抗WNT3(1∶200)及内参β-actin(1∶1500) 在4 ℃环境中孵育过夜。第2天加入经过碱性磷酸酶标记的二抗(1∶1000稀释),改为在37 ℃环境下孵育2 h,TBST缓冲液冲洗3次后用BCIP/NBT进行显色。结果通过Image J图像分析软件进行扫描测定图象灰度,得出目的及内参蛋白的光密度比值,即为目的蛋白表达水平的半定量指标。
1.3 统计学处理
实验结果通过均值±标准差表示,采用SPSS 11.5统计软件进行单因素方差分析,P<0.05表示差异有统计学意义。
2 结果
2.1 细胞生长状态观察
SD大鼠骨髓细胞中有大量悬浮细胞在第1次换液时会被丢弃,而在沉淀细胞中可见到单核细胞,呈现典型成纤维细胞样生长以梭型或多边型为主。细胞融汇瓶底至9/10大概需要10 d左右。
2.2 Real time RT-PCR结果
实验组与对照组两组细胞分别经诱导培养及诱导培养加用辛伐他汀干预12 h后,Real Time-PCR分析,实验组因子WNT3 mRNA比对照组的表达水平增高,两组差异有统计学意义(P<0.01)。经过36 h后,两组间因子WNT3 mRNA的表达水平差异有统计学意义(P<0.05)。详见表2。

表2 Real Time-PCR测定WNT3 MrnaTable 2 Real Time-PCR evaluation of WNT3 mRNA
注:实验组与对照组相比,△△P<0.01;△P<0.05。
2.3 Western blot
两组细胞在经诱导培养及加用辛伐他汀干预12 h后,因子WNT3的蛋白表达水平变化不大。36 h后WNT3蛋白表达水平实验组高于对照组,但是两组间的差异无统计学意义(P<0.05)。详见表3。
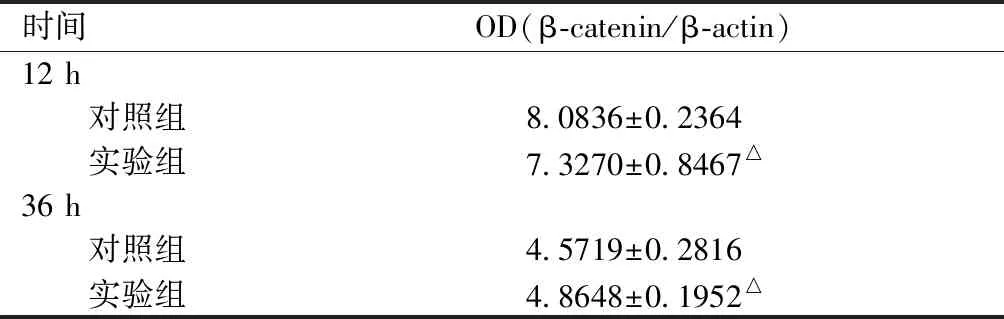
表 3 WNT3蛋白的表达Table 3 The expression level of WNT3 protein
注:实验组与对照组相比,△P>0.05。
3 讨论
通过本实验发现,体外短期应用辛伐他汀干预BMSCs 12 h及36 h后对其WNT信号通路主要相关因子在骨髓基质干细胞成骨分化早期有一定作用。
骨髓基质干细胞的多向分化潜能在多项研究中已有所发现,比如在诱导剂诱导下能够使BMSCs向成骨细胞分化。同时,BMSCs也可向软骨细胞分化[9]。不过在BMSCs向成骨细胞定向分化过程中参与的信号通路较多,其中主要的WNT信号通路相关因子参与调控并发挥重要作用[10-11]。
不论在骨组织工程学还是在骨折愈合及骨质疏松治疗学方面都具有重要价值的成骨细胞,如何使骨髓基质干细胞定向分化有着重要的研究价值。Wnt信号通路在MSCs体外分化所起的作用随仍存有争议,但大部分研究证实了其积极作用。而早期阶段的作用及其机制鲜有报道。有研究表明,在BMSCs向成骨细胞分化过程中经典的Wnt/β-catenin 信号通路发挥积极作用[12-13],而有的研究结果与之不一致而相冲突[14]。有的研究发现通过别的通路也能达到促进成骨细胞分化的作用[15]。尽管目前存有争议,但是都能一致认可BMSCs强大的成骨潜能,研究者认为BMSCs也参与了骨修复愈合过程,成为增加骨量、修复骨损伤最有前途的干细胞之一。
有关辛伐他汀在成骨分化早期所发挥作用的报道较少,涉及WNT信号通路早期的也鲜有报道。辛伐他汀在早期干预能促进骨髓基质干细胞向成骨细胞分化[16-17]。刘昊等[18]的研究也证实了这种分化潜能。通过本实验,两组间在辛伐他汀干预12 h及36 h后,WNT信号通路主要因子WNT3 mRNA表达水平实验组与对照组相比明显升高,而蛋白表达水平两组间未见明显差异。
本研究发现,经过辛伐他汀干预对SD大鼠骨髓基质干细胞的成骨分化有一定促进作用,涉及其中的重要信号通路主要因子的表达mRNA及蛋白并不一致,可能与观察时限有关,也可能有其他因素参与其中,其具体调节机制及其与其他不同因子或不同通路之间相互间作用有待进一步证实。
