Ano1在TGF-β1介导的肠道上皮细胞上皮间质转分化中的作用
2019-08-06樊晴伶王景杰窦维佳
樊晴伶,王景杰,窦维佳
(空军军医大学第二附属医院消化内科,西安 710038;#共同第一作者;*通讯作者,E-mail:weijia-dou@126.com)
肠道纤维化是克罗恩病(Crohn’s disease,CD)常见且严重的并发症之一。研究显示70%CD患者在病程中会发生纤维化并最终导致肠腔狭窄[1]。纤维化一旦发生,至少60%的患者在20年内需要手术治疗,然而术后狭窄亦有可能反复发作,严重影响患者生活质量[1]。肌成纤维细胞(myofibroblast,MF)是纤维化的主要效应细胞。肠道纤维化过程中,肠上皮细胞可通过上皮间质转分化(epithelial-mesenchymal transition,EMT)转换为MF[1,2],是MF的主要来源之一。EMT指上皮细胞失去其原有表型,获得向间质细胞转化的潜能。研究发现EMT参与肿瘤侵袭转移、组织纤维化、瘢痕修复等病理生理过程[3,4],该过程受多种生长因子及细胞因子调节。转化生长因子β1(transforming growth factor β1,TGF-β1)是EMT最有效的诱导因子[3]。研究发现在CD合并纤维化中TGF-β1表达明显升高[5]。因此,如何有效抑制TGF-β1诱导的EMT是预防和治疗肠道纤维化的关键。
Ano1即TMEM16A,是钙激活氯通道(calcium-activated chloride channels,CaCCs)成员之一,在维持正常生理功能方面有不可替代的重要作用[6]。近来大量研究发现,Ano1在多种肿瘤中表达上调,与肿瘤增殖、转移、预后不良密切相关[7]。在二硝基苯磺酸(DNBS)诱导肠道纤维化的大鼠模型中,肠道肌层中Ano1表达明显升高,其表达水平与Collagen I和弹力蛋白呈正相关[8],提示Ano1可能参与肠道纤维化过程,但其具体机制仍不十分明确。
本研究通过TGF-β1刺激大鼠小肠隐窝上皮细胞(IEC6),观察其细胞形态变化,并检测EMT相关标志物及Ano1表达情况。利用过表达慢病毒感染IEC6,上调细胞中Ano1表达后,再次以TGF-β1刺激感染细胞,观察细胞形态并检测EMT相关蛋白的表达情况,为进一步动物实验及临床应用提供实验依据和理论依据。
1 材料与方法
1.1 主要试剂和仪器
Ano1过表达慢病毒(由上海吉凯基因技术有限公司合成),M-MLV逆转录酶(美国Promega公司),胎牛血清FBS(美国Invitrogen公司),TGF-β1蛋白(英国Abcam公司),兔抗α-SMA单克隆抗体、兔抗vimentin单克隆抗体、小鼠抗E-cadherin单克隆抗体(英国Abcam公司),小鼠抗Fibronectin单克隆抗体(美国R&D公司),兔抗Ano1多克隆抗体(美国Proteintech公司)、GAPDH抗体、羊抗兔二抗、羊抗小鼠二抗(美国Santa Cruz公司),DMEM培养液(Hyclone公司),311型隔热式CO2培养箱(Thermo Fisher Scientific),5417R型低速低温离心机(Eppendorf),XDS-100型倒置显微镜(上海蔡康光学仪器有限公司),IX71型荧光显微镜(日本Olympus公司)。
1.2 方法
1.2.1 细胞培养 大鼠小肠隐窝上皮细胞(IEC6)购自中国科学院上海细胞库,DMEM+10%FBS于37 ℃、5% CO2条件下常规培养。
1.2.2 慢病毒转染 取对数生长期IEC6,制成浓度为1×104个/ml的细胞悬液,接种1 ml细胞悬液于12孔培养板中。细胞贴壁生长至20%融合时,吸取上清,加入2 μl Ano1过表达慢病毒(2×10-8TU/ml),16 h后观察细胞状态,替换为常规培养基继续培养,构建Ano1过表达IEC6细胞系(IEC6-Ano1),以空载体为阴性对照(IEC6-NC)。
1.2.3 实验分组及处理 明确TGF-β1对IEC6细胞EMT过程及Ano1表达的影响,实验分为两组:①control组(正常对照组):IEC6加入无血清DMEM培养基,培养72 h;②TGF-β1组:IEC6加入无血清DMEM培养基,培养过夜,第2天加入10 ng/ml TGF-β1培养72 h,倒置相差显微镜观察不同组细胞形态,并弃上清收细胞,用于后续实验。
进一步探讨Ano1在TGF-β1诱导IEC6细胞EMT中的作用,实验分为三组:①control组(正常对照组):IEC6加入无血清DMEM培养基,培养72 h;②IEC6-NC组:将IEC6-NC细胞加入无血清DMEM培养基,培养72 h;③IEC6-Ano1+TGF-β1组:IEC6-Ano1细胞加入无血清DMEM培养基,培养过夜,第2天加入10 ng/ml TGF-β1培养72 h,荧光显微镜观察不同组细胞形态,并弃上清收细胞,用于后续试验。
1.2.4 real time-PCR检测Ano1 mRNA表达水平 使用Trizol法提取细胞总RNA,紫外分光光度法定量,并统一各组RNA浓度。取2 μl总RNA,在M-MLV逆转录酶作用下,逆转录合成cDNA,以合成的cDNA为模板进行PCR检测,反应条件:95 ℃,3 min变性,95 ℃ 12 s,62 ℃ 40 s,共40个循环。采用2-ΔΔCt分析mRNA相对表达水平,每个样本设置3个复孔,实验重复3次。以GAPDH为内参,所有引物均由生工公司合成,Ano1及GAPDH引物序列如下。Ano1:上游5′-TCGAGGAAGAGGAGGTGAGTAG-3′,下游5′-GATGTTGGACCGCACAGATG-3′;GAPDH:上游5′-GACAGTCAGCCGCATCTTCT-3′,下游5′-GCGCCCAATACGACCAAATC-3′。
1.2.5 Western blot检测Ano1、E-cadherin、Vimentin、Fibronectin及α-SMA蛋白表达水平 收集对数生长期细胞,PBS冲洗3次,加入100 μl细胞裂解液充分裂解,4 ℃、12 000g,离心15 min,取上清BCA法测定蛋白浓度。根据蛋白定量结果取15-20 μl蛋白样品加于10% SDS-PAGE中。电泳结束后将蛋白转移到PVDF膜,含5%脱脂奶粉的TBST室温封闭1 h,分别加入抗Ano1(1 ∶500稀释)、E-cadherin(1 ∶1 000稀释)、Vimentin(1 ∶1 000稀释)、Fibronectin(1 ∶500稀释)及α-SMA(1 ∶1 000稀释)抗体,4 ℃过夜。TBST洗膜4次,每次8 min,加入二抗(1 ∶5 000稀释)室温孵育2 h,TBST洗膜4次,每次8 min,ECL显色,采集图像后用Image J图像软件系统进行分析。
1.3 统计方法
所有数据均采用均数±标准差表示,用SPSS16.0软件进行统计,两组间差异比较采用t检验,多组间差异比较采用单因素方差分析,P<0.05认为差异有统计学意义。
2 结果
2.1 TGF-β1诱导IEC6发生EMT
以10 ng/ml TGF-β1刺激IEC6,72 h后观察发现,与正常对照组相比,IEC6形态发生明显改变,由纺锤形变为长梭形或多角形,且细胞极性消失(见图1)。Western blot结果显示,上皮相关分子标志物E-cadherin蛋白表达水平降低,而间质相关分子标志物vimentin、Fibronectin及α-SMA蛋白表达水平明细升高,差异有统计学意义(P<0.001,见图2),说明TGF-β1可诱导IEC6发生EMT。
2.2 TGF-β1抑制IEC6中Ano1表达
RT-PCR结果显示,与正常对照相比,TGF-β1刺激后IEC6中Ano1 mRNA表达水平降低(P<0.05,见图3);Western blot结果证实Ano1蛋白表达水平亦显著降低(P<0.05,见图2),说明TGF-β1可抑制IEC6中Ano1的表达。

A.control组,×10;B.control组,×40;C.TGF-β1组,×10;D.TGF-β1组,×40图1 TGF-β1刺激IEC6细胞形态变化Figure 1 Effect of TGF-β1 on IEC-6 morphology
2.3 TGF-β1对IEC6-Ano1细胞形态的影响
利用Ano1过表达慢病毒感染IEC6,上调Ano1表达,以空载体质粒为阴性对照组,观察发现与control组相比,阴性对照组(IEC6-NC)细胞形态无明显改变,而以10 ng/ml TGF-β1刺激过表达Ano1的IEC6(IEC6-Ano1),细胞形态亦无明显改变,仅个别细胞呈多角化趋势(见图4)。

与control比较,*P<0.05,**P<0.001图2 TGF-β1刺激IEC6后EMT相关蛋白及Ano1的表达Figure 2 Expression level of EMT-related proteins and Ano1 in IEC6 cells stimulated by TGF-β1
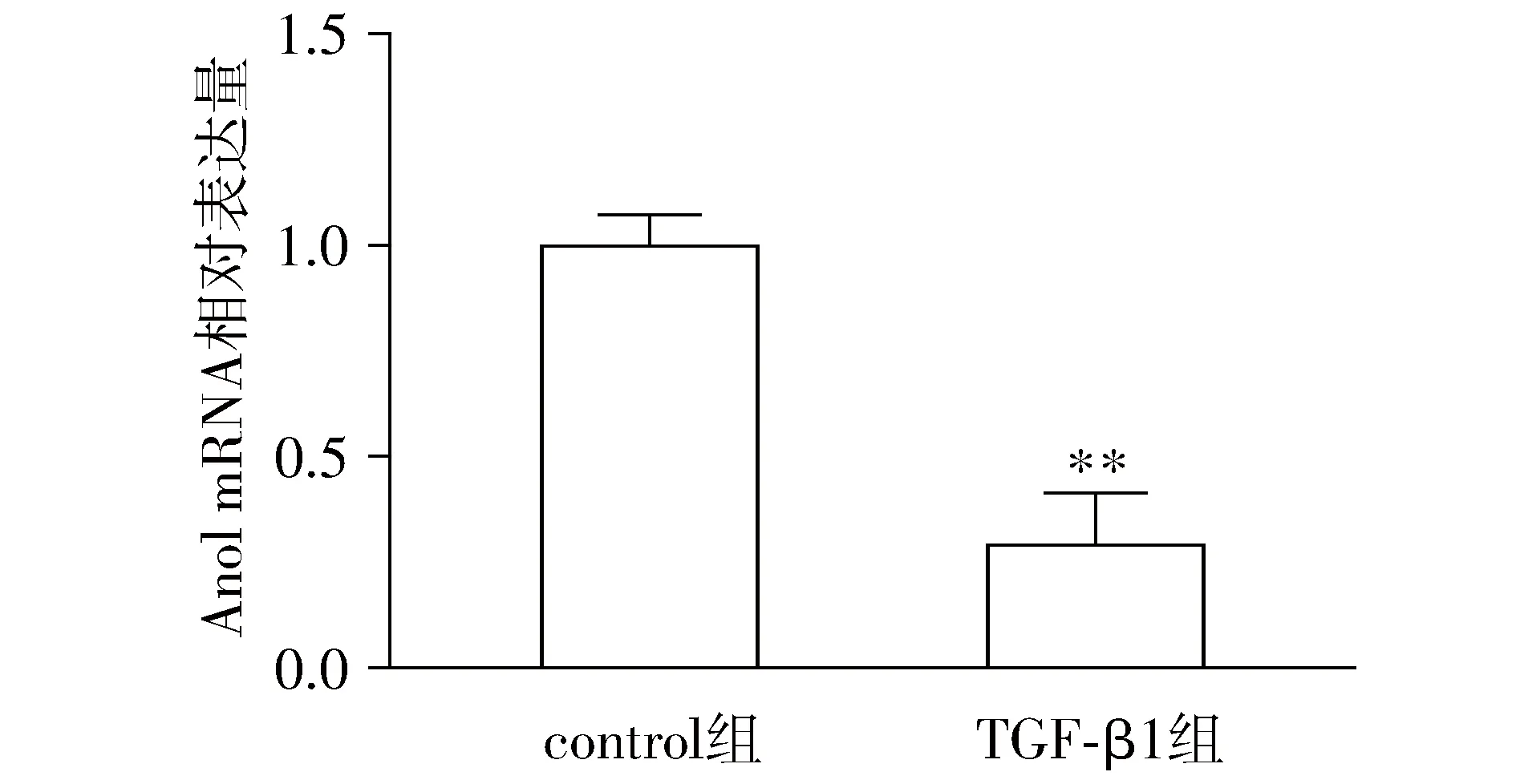
与control组比较,**P<0.05图3 TGF-β1刺激IEC6后72 h Ano1 mRNA的水平Figure 3 Expression level of Ano1 mRNA in IEC6 cells stimulated by TGF-β1 for 72 h
2.4 TGF-β1对IEC6-Ano1细胞EMT相关分子标志物的影响
以10 ng/ml TGF-β1刺激IEC6-Ano1细胞72 h后,Western blot结果显示,E-cadherin、Vimentin、Fribronectin及α-SMA蛋白表达水平较control组及IEC6-NC组无明显差异(见图5),说明在IEC6中上调Ano1可抑制TGF-β1介导的EMT发生。
3 讨论
肠道纤维化是克罗恩病常见且严重的并发症,其发病机制复杂,目前尚不十分明确。传统观念认为,肠道反复慢性炎症刺激可激活肌成纤维细胞,在肠道纤维化中起重要作用[9]。近年来研究发现,纤维化与局部炎症的严重程度无明显相关性[10]。纤维化一旦发生,即使肠道炎症得到控制也无法预防或逆转纤维化进展[11],因此,阐明肠道纤维化发生发展机制,寻找可能的预防及治疗方法,具有重要意义。

图4 TGF-β1刺激IEC6-Ano1后细胞形态变化Figure 4 Effect of TGF-β1 on IEC6-Ano1 morphology

图5 TGF-β1刺激IEC6-Ano1后EMT相关蛋白的表达Figure 5 Expression level of EMT-related proteins in IEC6-Ano1 cells stimulated by TGF-β1
EMT是指具有极性的上皮细胞发生间质细胞表型的转变,在EMT过程中,上皮细胞失去其原有细胞形态、E-cadherin等蛋白表达降低,获得间质细胞长梭形或多角形形态,且Vimentin、α-SMA等间质蛋白表达增加。研究表明,EMT不仅参与胚胎生成、器官发育、组织再生和伤口愈合等过程[12],同时在肾脏、肺脏、肠道等多种器官及组织纤维化中发挥重要作用[3]。Scharl等[13]在克罗恩病患者肠道纤维化部位发现,Fribronectin及α-SMA等EMT相关蛋白表达增加。Flier等[14]通过大鼠直肠内注射三硝基苯磺酸溶液(TNBS)模拟克罗恩病肠道炎症和纤维化过程,发现约1/3肌成纤维细胞来源于肠上皮细胞,提示EMT参与肠道纤维化。另有研究发现在TNBS诱导的大鼠结肠纤维化模型中,miR-200b可以抑制结肠上皮细胞EMT过程,从而缓解肠道纤维化程度[15]。因此,抑制EMT的发生发展可作为治疗肠道纤维化的潜在靶点之一。
TGF-β1是纤维化过程的重要调节因子。TGF-β1促使细胞外基质沉积,成纤维细胞增生,抑制细胞外基质降解,在慢性肾病、肝脏、肺纤维化等病理生理过程中发挥重要作用[16,17]。另有研究发现在CD合并纤维化组织中,TGF-β1表达显著升高[5],提示TGF-β1亦可参与肠道纤维化过程。大量研究显示TGF-β1通过上调MMP9、PDGF等细胞因子,激活RTK、Notch等信号通路,诱导细胞EMT过程,最终导致纤维化的发生[18]。本研究我们利用TGF-β1刺激IEC6细胞观察发现,刺激后IEC6细胞形态发生间质样改变,上皮相关分子标志物E-cadherin蛋白表达水平显著降低,而间质相关分子标志物Vimentin、Fibronectin及α-SMA蛋白表达水平显著升高,提示TGF-β1可诱导IEC6细胞发生EMT。
Ano1是一种跨膜蛋白,表达于分泌上皮、血管平滑肌细胞、ICC、感觉神经元等多种正常组织及细胞,介导不同器官和组织的生理功能和生物学行为[19]。近年来研究发现,Ano1在头颈部鳞癌、食管癌、乳腺癌、前列腺癌和胃肠间质瘤等多种肿瘤中表达上调,在增殖、侵袭、转移等肿瘤恶性生物学行为中发挥重要作用[4,6]。另外,Gao等[20]在心梗诱发心脏纤维化小鼠模型中发现,心脏成纤维细胞中Ano1表达升高,心内注射过表达Ano1腺病毒继续上调Ano1表达,心脏纤维化程度却有所缓解,同时发现Ano1可抑制TGF-β/Smad3信号通路。而Sun等[21]发现在囊性纤维化中,TGF-β可抑制结肠上皮细胞T84中Ano1的表达。为了揭示Ano1在肠道纤维化中的作用,我们利用TGF-β1诱导IEC6发生EMT,发现TGF-β1可显著抑制IEC6中Ano1的表达,提示Ano1可能参与TGF-β1诱导的肠上皮细胞EMT过程。进一步研究中我们应用慢病毒系统上调IEC6中Ano1表达,再次给予TGF-β1刺激,发现过表达Ano1后IEC6的EMT过程被抑制,表现为细胞形态无明显改变,且E-cadherin、Vimentin、Fribronectin、α-SMA蛋白表达水平无明显变化。
综上所述,我们的研究结果表明Ano1参与TGF-β1介导的IEC6细胞EMT过程,在IEC6细胞中上调Ano1表达,可以抑制TGF-β1诱导的EMT。
