珍稀特有植物华顶杜鹃的种群结构和种间联结
2019-08-05蔡鑫陈波陈锋陈伟杰陈子林王盼陈珍慧金孝锋
蔡鑫,陈波*,陈锋,陈伟杰,陈子林,王盼,陈珍慧,金孝锋*
(1.杭州师范大学生命与环境科学学院,浙江杭州310036;2.浙江省森林资源监测中心,浙江杭州310020;3.浙江省大盘山国家级自然保护区管理局,浙江磐安322300)
种群结构是植物群落的基本属性,也是认识群落组成、变化和发展的基础。种群结构除了能够分析当下的种群状况,还能了解过去和预测未来的种群状态与动态,从而反映群落的演替趋势[1-4]。植物群落中物种在空间分布上的相互关联性,即种间联结,通常是由群落生境差异影响物种的分布而引起。这种联结性是各物种在不同生境中相互影响形成的有机联系的反应。种间联结研究对维护植物群落的稳定性、保护群落的多样性具有重要意义,且对特定物种的保护也有比较重要的作用[5-7]。因此,将种群结构和群落种间联结起来分析,可以更好地阐明物种的濒危机制和群落的竞争状况,预测群落的演替趋势。
华顶杜鹃(Rhododendron huadingenseB.Y.Ding&Y.Y.Fang)是杜鹃花科(Ericaceae)杜鹃花属(Rhododendron)植物,特产于浙江省,是浙江省省级重点保护植物,曾报道只分布在模式产地天台华顶山[8-9],近几年又陆续在浙江省大盘山国家级自然保护区、磐安高姥山、余姚四明山、金华北山被发现[10-12]。作为浙江省珍稀濒危植物,其毛被特征、花序类型、果实形状和种子大小在杜鹃花属中颇为特殊,在系统发育研究中具有重要价值[13],其花大艳丽,先叶开放,极具观赏价值。已有的关于华顶杜鹃形态特征、系统分类位置[8,13]和少量分布区群落特征[10]的报道显示,华顶杜鹃所面临的竞争压力较大,株数随基径级增大逐渐减少,呈空间聚集分布格局[9,14];所处生境温热多湿,群落植物组成丰富,群落异质性大[10-11]。目前,尚未见对华顶杜鹃的种群结构和群落主要种群间的联结性研究的报道。本研究在对华顶杜鹃浙江省所有分布区野外调查的基础上,从群落学、种群生态学的角度,分析了种群结构和种间联结情况,以揭示华顶杜鹃群落不同结构层次主要物种的种间关系,探讨其濒危机制,为其有效保护、生境创造和种群恢复提供科学依据。
1 材料与方法
1.1 野外调查
基于已有研究和野外踏查,选取分布于华顶山(HDS)、大盘山(DPS)、高姥山(GLS)、四明山(SMS)和北山(BS)等地的华顶杜鹃典型群落,研究地自然概况可见相关文献[11,15-17]。根据华顶杜鹃现有分布情况,共设置10个绝对距离为20 m×20 m的样地(各样地的基本概况见表1)。将每个样地划分为16个5 m×5 m的样方,调查时记录每个样方中乔木层(胸径>3 cm)的种类、胸径、树高、冠幅等;在每个5 m×5 m样方的左下角设2 m×2 m的小样方,用以调查灌木层(胸径<3 cm),调查时记录种名、株丛数、平均高度、盖度等;并记录华顶杜鹃每个个体的基径(其树干不高,在1.3 m以下分枝较多)、高度、冠幅,用以种群年龄结构分析。同时,以大小级代替年龄,参考浙江百山祖木本植物径级划分法[18],将华顶杜鹃基径<1 cm且树高<33 cm的个体划分为幼苗(等级Ⅰ),基径<1 cm且树高>33 cm的划分为幼树(等级Ⅱ),基径>1 cm的划分为7个等级(等级Ⅲ~Ⅸ)。
1.2 数据处理与分析方法
1.2.1 优势度计算
用重要值(IV)表示物种在群落中的优势程度,计算公式为

式中,RP表示相对显著度,RA表示相对多度,RF表示相对频度[10]。

表1 华顶杜鹃样方概况表Table 1 The environmental conditions of the studied 10 plots
1.2.2 群落相似性
用Sorensen相似性指数(SC)表示群落之间的相似性,计算公式为
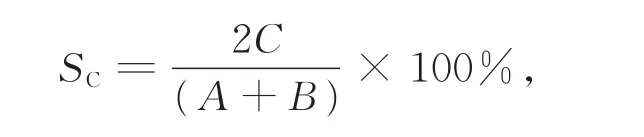
A、B分别为甲、乙两样地的植物种数;C为两样地共有的植物种数[19]。
1.2.3 总体关联性检验
用SCHULTER[20]提出的方差比率法(VR)检测多物种间的关联性,统计量公式为:
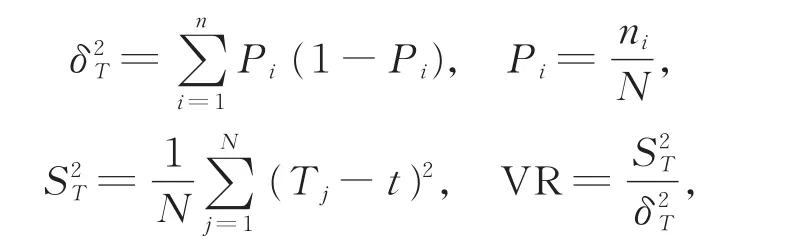
其中,S为总物种数,N为总样地数,Tj为样地j内出现的研究物种总数,ni为物种i出现的样地数,t为每个样地中种的平均数,t=(T1+T2+…+Tn)/N,Pi为物种i出现的样地数占总样地数的比例,S2T为所有样地物种数的方差,δ2T为所有物种出现频度的方差。当VR>1时,表示物种表现出正关联;当VR<1时,表示物种间出现负关联;当VR=1时,即符合所有种间无关联的零假设。VR偏离1的显著性用统计量W来检验:W=VR×N。若种间无关联,则W落在(χ20.95,N,χ20.05,N)的概率有 90%[21]。
1.2.4 种间联结测度方法
根据样地调查资料,选取10个样地内的优势物种(重要值>5%),建立2 m×2 m联列表,分别计算a、b、c、d值。其中,a为2个种都出现的样地数;b、c分别为仅有种A或种B出现的样地数;d为2个种均未出现的样地数;N为总取样单位。当某个种的频度为100%时,b、d值加权为1。共同出现百分率PC、联结系数 AC、Pearson积矩相关系数(rp)和 Spearman秩相关系数(rs)的计算公式和检验方法[21]如下:
(1)PC又称Jaccard指数(JI),用来检测物种间的正联结程度,计算公式为

其值域为[0,1],越趋近于1,表明该种对的正联结越紧密;若PC=0,表明种对无关联。
(2)联结系数AC用来说明种间的联结程度,计算公式为
若ad≥bc,则
若bc>ad且d≥a,则
若bc>ad且d<a,则
AC值域为[-1,1],AC值为0,说明2个物种完全独立;AC值越趋近于1,说明2个物种间正联结越强;AC值越趋近于—1,说明2个物种间负联结越强。
(3)Pearson积矩相关系数(rp)和Speraman秩相关系数(rs)是反映2个物种种间协变线性关系的重要指标,可用来定量分析2个物种间的线性关系。
Pearson积矩相关系数:
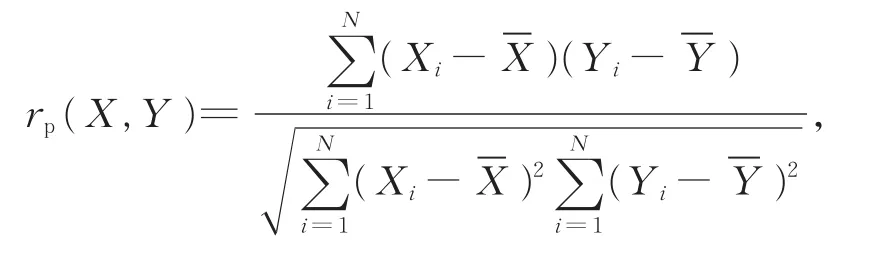
Spearman秩相关系数:
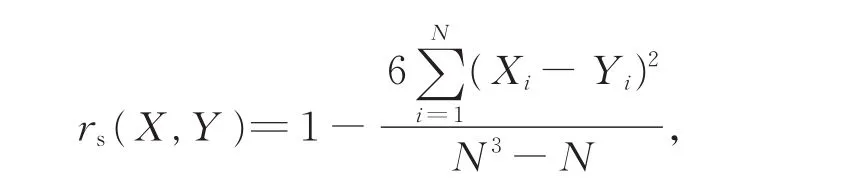
式中,N为总样地数,Xi和Yi分别表示种X、Y在样地i中的重要值。
2 结果与分析
2.1 群落相似性
华顶杜鹃各样地间的Sorensen相似性系数SC值集中在 0.179~0.585,平均值为 0.333(见表 2)。SC值大于 0.5的有 4对,为 DPSc-DPSd(SC=0.585)、DPSb-DPSc(SC=0.522)、DPSb-DPSd(SC=0.514)、SMSa-SMSb(SC=0.525)。HDSa-BS的SC值最小,为0.179。

表2 华顶杜鹃群落间Sorensen相似性系数Table 2 Sorensen's similarity coefficient between different communities of Rhododendron huadingense
2.2 种群结构分析
华顶杜鹃在10个样地中共计个体451株,幼苗和幼树分别占总株数的8.42%和17.07%,Ⅵ级及以上的占比为46.5%。总体上,华顶杜鹃大径级植株所占比例较高,Ⅱ级及以下所占比例较低,幼苗和幼树储备较少,种群正趋于衰退期。
各样地的径级结构不连续(见图1)。DPS的4个样地幼苗和幼树相对较多,中等径级植株缺失。HDSa、SMSb、GLS样地的华顶杜鹃以中等径级植株为主,大径级植株较少或缺失,HDSb、SMSa样地的幼苗和幼树缺失,均为中等径级或大径级植株。
2.3 总体联结性
根据华顶杜鹃群落中的优势种,最终确定参与群落相关分析的物种,乔木层22种、灌木层24种。乔木层和灌木层的总体关联性由表3所示,可见乔木层表现出不显著的正关联,灌木层表现出不显著的负关联。
2.4 华顶杜鹃群落种间的关联性
通过分析共同出现百分率PC、联结系数AC对华顶杜鹃群落种间的关联性,依据乔木层22个树种231个种对、灌木层24个树种276个种对共同出现的百分率和联结系数来测定(见图2和图3)。
从图2(a)可见,华顶杜鹃乔木层231个种对共同出现的百分率检测结果显示,PC≥0.6的种对有16个,其中PC=1的有4对,分别为16-18(华中樱Cerasus conradinae-灰叶安息香Styrax calvescens)、3-17(马尾松Pinus massoniana-黄檀Dalbergia hupeana)、3-19(马尾松-山橿Lindera reflexa)、17-19(黄檀-山橿),所占比例为1.73%;PC值显示,相互间无关联的种对(P=0)有101个,即这些种对在样方中从未同时出现过,所占比例为43.7%;关联度在0~0.6的种对有114个。乔木层与华顶杜鹃种群联结程度较高的种群有化香Platycarya strobilacea,联结程度弱的有马尾松、天台鹅耳枥Carpinus tientaiensis、野桐Mallotus japonicus等。

图1 各样方中华顶杜鹃的径级结构Fig.1 The diameter class structure of Rhododendron huadingense in 10 plots
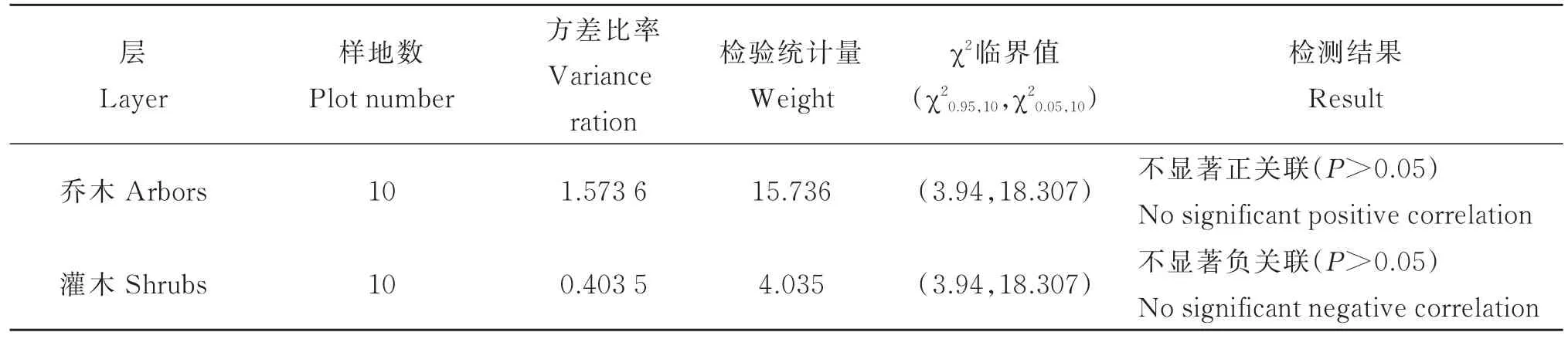
表3 群落各层次主要种间的总体关联性Table 3 Overall interspecific associations among dominant plant populations of different layers
如图3(a)所示,由华顶杜鹃灌木层276个种对的PC值计算显示,PC=1的种对有4个,如14-1(华顶杜鹃-微 毛 柃Euryahebeclados)、15-25( 满 山 红Rhododendron mariesii-算盘子Glochidion puberum)等。0.6≤PC<1的有10个,如14-9(华顶杜鹃-映山红Rhododendron simsii)、3-5(茅栗Castanea seguinii-山橿)等;0<PC<0.6的种对有160个;PC=0的种对有82个。
图2(b)、图3(b)表明,乔木层和灌木层种对联结系数AC值域为[—0.6,1],与PC值整体表现一致,乔木层中种对16-18(华中樱-灰叶安息香)、3-17-19(马尾松-黄檀-山橿),灌木层中14-1(华顶杜鹃-微毛柃)、15-25(满山红-算盘子)的AC值为1,表明这些种对完全联结;乔木层中AC=—1的种对有9对,灌木层有35对;乔木层和灌木层中0.6≤|AC|<1的种对分别为9对和8对;其余种对|AC|<0.6,表明物种间呈不显著的正关联或负关联。
2.5 Pearson积矩相关系数rp和Spearman秩相关系数rs
Pearson积矩相关系数和Spearman秩相关系数检验结果如表4所示。Pearson系数结果表明,乔木层231个种对中,正相关的种对数为61个,占26.4%,其中11对呈极显著正相关,7对呈显著正相关;负相关的种对数有171对,占73.6%。灌木层276个种对中,呈正相关的有85对,占30.8%,其中11对呈极显著正相关,10对呈显著正相关;负相关的种对有181对,种对1-3(微毛柃-茅栗)呈极显著负相关。
Spearman秩相关系数检验结果显示,乔木层231个种对中,正相关的为93对,其中,极显著正相关的有6对,显著相关的有8对;负相关的有138对,占57.7%,无显著负相关的种对。灌木层呈正相关的种对有127对,其中,6对呈极显著正相关,9对呈显著正相关;149个种对呈负相关,种对1-3(微毛柃-茅栗)呈极显著负相关,种对5-11(山橿-山胡椒Lindera glauca)、5-16(山橿-中国绣球Hydrangea chinensis)、14-21(华 顶 杜 鹃 -蜡 瓣 花Corylopsis sinensis)呈显著负相关。

图2 华顶杜鹃样地乔木层半矩阵图Fig.2 Half-matrix diagram of Rhododendron huadingense communities(arbors layer)
3 讨 论
现已知的华顶杜鹃分布在海拔800~1 150 m,为中高海拔狭域分布种。根据已发现华顶杜鹃的分布情况,可知其大多分布在光线较强的阳坡,且分布在较贫瘠的土壤[22]。地形变化因素引起的生境异质性致使群落间物种组成变化明显,从而影响植物种群结构及物种多样性[23-24]。华顶杜鹃群落物种组成在不同产地的差异较大,同一产地的差异较小,除地理距离较近的DPS的3个样地、SMS的2个样地SC值较高外,其余样地SC值均不高。
种群年龄结构分析是解释种群结构现状和更新策略的重要途径之一。华顶杜鹃的10个样地中,种群普遍较小,各样地均处于不同程度的衰退阶段(见图1),随着时间的推移,种群将趋于老化,且各样地龄级结构不连续,相对而言,大盘山的华顶杜鹃种群衰退程度较低,更具有发展潜力。导致华顶杜鹃天然更新能力差的主要原因可能有三方面:一是华顶杜鹃分布在阳坡,土壤有机质积累少、土壤干燥且较贫瘠;二是人类活动对其干扰大,华顶山的样地分别位于停车场和雷达站附近,高姥山的样地靠近路边,四明山和北山的样地位于林场中,早年砍伐严重;三是华顶杜鹃观赏价值高易被采挖。大盘山4个样地均处于保护区,以大径级植株为主,幼苗和幼树次之,中等径级的植株缺失。可能是早年人为砍伐等干扰导致华顶杜鹃数量下降,种群结构趋于老龄化。保护区的建立,加强了对华顶杜鹃的保护,干扰因素减少,为幼苗的更新和所处群落的稳定创造了条件。小种群濒危物种的保护是我国当前生物多样性保护的重要内容之一,如何有效保护这些物种是一项艰巨的任务[25]。对华顶杜鹃的保护,关键是对其所处森林生态系统的保护。

图3 华顶杜鹃样地灌木层半矩阵图Fig.3 Half-matrix diagram of Rhododendron huadingense communities(shrubs layer)
种间联结表示物种之间相互关系的静态描述,包括空间分布关系,也包含物种之间的功能关系,正或负的种间联结对群落的演替起一定的作用,分析群落的种间联结有利于认识群落的演替方向[26-27]。华顶杜鹃各样地的种间总体相关性、种间联结性(PC值和AC值)、种间相关性(Pearson相关系数和Spearman秩相关系数)检验结果(见表3、图2和图3)基本一致,总体表现为:乔木层为不显著正关联,灌木层为不显著负关联,绝大多数种对间联结性不显著,独立性较强,且未形成稳定的搭配关系。原因可能有两方面:一是植物在个体发育和群落演替过程中,受到各方面的影响,包括土壤、水分等环境因素,不同层次、同层次间的植物也要进行激烈的竞争;二是不同物种对环境的要求不同,分别占有不同的生态位。灌木层的华顶杜鹃只与蜡瓣花呈显著负相关,可能因两者皆为阳生植物,且群落乔木层盖度较大(60%~85%),可能由于两者之间生态位的重叠较多,进而产生对光资源的竞争[28]。

表4 华顶杜鹃群落的Pearson积矩相关系数和Spearman秩相关系数检验结果Table 4 The test between species in communities of Rhododendron huadingense,produced by Pearson's correlation coefficient and Spearman's rank correlation
群落成熟度愈高,物种间应存在较强的正关联,以求多物种稳定共存,处于环境资源利用的最佳状态[29-30]。华顶杜鹃群落物种间负关联明显多于正关联,可能由于群落中物种的生态习性差异较大,在共存时无法实现对资源的最佳分配和利用,群落处于不稳定阶段。但种间关联性只能解释表面现象,揭示的是种间竞争的结果或群落的现状,难以深入阐明群落种间关系的内在原因[31]。
对华顶杜鹃的种群结构和群落内种间关系研究的结果表明,2种方法的研究结论有较大的相似性,同时互为补充,在实践中可采取以下措施保护与管理华顶杜鹃野生资源,抑制其种群的衰退趋势:首先,在加强对华顶杜鹃个体保护的同时,要重视对群落及其生境的保护。当群落生境受到人为或其他干扰时,易改变群落内的水热条件,使物种组成发生变化,从而改变物种之间的相互关系。其次,为促进华顶杜鹃自然更新,应适当人为干预群落中对华顶杜鹃有负面作用的物种。
