茶树表没食子儿茶素-3-O-(3-O-甲基)没食子酸酯研究进展
2019-08-02孙威江邓婷婷
谢 凤,孙威江,邓婷婷
1福建农林大学园艺学院,福州 350002;2福建省安溪县现代农业产业园协同创新中心,泉州 362400;3福建农林大学安溪茶学院,泉州 362400;4福建省茶产业工程技术研究中心,福州 350002
1982年,EGCG3"Me((-)-Epigallocatechin-3-O-(3-O-methyl) gallate)首次由日本科学家从绿茶中分离,EGCG3"Me在抗肿瘤、降血压、增加肠道有益菌等保健功效均优于表儿茶素没食子酸酯(EGCG),特别是EGCG3"Me是目前发现抗过敏作用最为显著的物质成分[1]。过敏性疾病具有遗传倾向,一些过敏性疾病仍然“流行”。由花粉、尘螨、真菌和宠物等致敏因素引起的过敏性鼻炎和哮喘最常见,季节性过敏性鼻炎如果不治疗,25%至38%会患哮喘,最终成为常年性哮喘,肺气肿和肺心病。据世界卫生组织估计,全球1.5亿哮喘患者中,50%的成人及至少80%的儿童是由过敏因素引起的,并逐年上升,每年有超过18万人死于哮喘。该类疾病已列为“21世纪重点研究和预防的疾病”[2]。
EGCG3"Me具有比EGCG更好的抗过敏效果[3],但目前已筛选出的高EGCG3"Me含量茶树资源较少,天然资源非常有限。绿色、高效的体外合成技术方法目前仍未有突破,对EGCG3"Me合成的研究多停在体外研究实验。茶树体内EGCG3"Me含量的影响因素多,EGCG3"Me的体内代谢途径也尚未研究清楚。目前国外对EGCG3"Me的研究多以保健功效研究为主,国内则集中在高EGCG3"Me种质资源筛选、茶树EGCG3"Me影响因素探究、EGCG3"Me分离合成方法的建立等方面。
1 EGCG3"Me理化性质及制备方法
1.1 EGCG3"Me理化性质
EGCG3"Me是茶树体内检测到的甲基化EGCG之一,与其它甲基化EGCG相比更常见且含量更高。甲基化反应广泛存在于生物体中,也称为醚化,EGCG苯环上的8个酚羟基部分或全部转化为甲基形成的一系列衍生物为甲基化EGCG[4]。EGCG3"Me是甲基取代了EGCG的D环没食子酰基3"的位置羟基,如图1。
EGCG3"Me是易溶于水和有机溶剂的白色粉末状物质。甲基取代不同位置的酚羟基后其抗氧化性能也发生变化[5],EGCG3"Me清除DPPH活性和总抗氧化活性明显低于EGCG[6]。但相对于EGCG表现出稳定性增加、脂溶性上升、更易吸收、生物活性更强等特征,口服吸收率比EGCG高9倍[6-9]。此外,EGCG3"Me还被证明口服后具有高药时曲线下面积(AUC),即高药物的生物利用度,这与其高生物活性和分布体积小有关[10]。
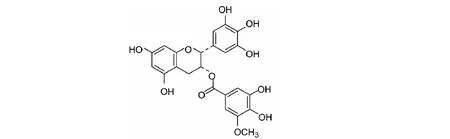
图1 EGCG3"Me分子结构式Fig.1 Molecular structural of EGCG3"Me
1.2 EGCG3"Me体外分离制备方法
甲基化EGCG分离方法包括制备型高效液相色谱(pHPLC)、Sephadex LH-20凝胶柱层析、高速逆流色谱(HSCCC)、Toyopearl HW-40S中压柱层析等。孙业良等[11]用制备型高效液相色谱分离、收集,经薄层色谱检测(TLC)检测和高效液相色谱(HPLC)分析,收集的组分釆用Sephadex LH-20进一步去除色素得EGCG3"Me纯品。龚正礼等[12]发明了利用茶叶制备甲基化儿茶素的方法。茶叶粉末提取萃取经粗聚酰胺和细聚酰胺树脂分离纯化得到甲基化儿茶素组分,经高速逆流色谱进一步分离得到高纯度的EGCG3"Me。周蓓等[13]以日本的茶叶为材料,处理后得到茶多酚粗品,粗品经Toyopearl HW-40S柱层析分离得到2种纯度较高的组分,一种为EGCG3"Me。
1.3 EGCG3"Me体外合成制备方法
目前,EGCG3"Me体外合成主要是用化学合成方法或生物合成方法修饰EGCG的甲基化结构。化学合成方法包括重氮甲烷合成法、碘甲烷合成法、硫酸二甲酯合成法、苄基保护基团合成法、硝基苯磺酰基保护基团合成法等,具有强目的性和高产率,但目前使用的大多数甲基化试剂是高毒性物质,并且非常需要纯化方法和药理学方面的后期处理方法。化学反应过程带来许多副产物,使分离纯化变得困难。生物学合成方法主要是酶法合成,酶是生物体内的一种催化剂,它能绿色、高效、有选择地在温和的条件下催化各类反应。但酶法合成中酶的提取技术尚不成熟,酶促反应产物不确定,如费冬梅[14]进行酶促反应实验一共生成了10种甲基化EGCG,其中包括4种一甲基化的EGCG、4种二甲基化的EGCG、2种三甲基化的EGCG,它们难以分离和纯化,再加上酶的不稳定性,酶法合成的成本高[15]。
2 EGCG3"Me保健功效
EGCG3"Me主要具有抗过敏、改善肠道、预防糖尿病、防肥胖、降血压、保护肝脏等保健功效,在体外和体内的临床实验中已经证实其在多方面表现出优异的生物学功能,具有进行深加工的潜力和价值,可开发专一功效为药用产品,也可开发综合功能为保健产品。
2.1 抗过敏
过敏性疾病,也称变态反应性疾病,是由患者过度敏感,在血液中产生特异性免疫球蛋白E抗体(IgE)导致过敏[2]。EGCG3"Me可有效控制花粉过敏反应症状,一是,茶叶中的EGCG3"Me可以阻隔IgE受体与过敏原,进而起到抗过敏的作用[16];二是,EGCG3"Me可以抑制高亲和力IgE Fc 受体(FcεRI)引发的神经刺激及直接抑制PS/TK活性,阻隔外界刺激作用于嗜碱细胞,从而抑制组胺释放达到抗过敏效果;三是,EGCG3"Me能抑制肥大细胞酪氨酸蛋白激酶(PTK)活性,可有效地阻断肥大细胞释放组胺[17,18]。还有研究表明[19],EGCG3"Me抑制小鼠皮肤被动反应的效果是EGCG的两倍。
2.2 改善肠道
EGCG3"Me对于改善人体肠道环境、维护人体肠道平衡具有重要作用[20]。EGCG3"Me能促进肠道有益细菌(双歧杆菌和乳酸菌)的增殖,并抑制有害细菌(梭状菌和拟杆菌)的增殖,EGCG3"Me的纯度越高,作用的效果越为明显[21-23],但对肠道总菌群数量无影响。张鑫等[21]进行体外厌氧粪样混合培养与荧光原位杂交技术实验,得出纯度90%EGCG3"Me样品的益生指数(PI)最高,为2.68,50%EGCG3"Me、30% EGCG3"Me和90% EGCG的样品PI值依次为2.25、2.14和1.86,而对照组的PI值仅为0.12。
2.3 预防糖尿病
糖尿病是一组代谢性疾病,其特征在于胰岛素分泌缺陷(Ⅰ型)或胰岛素作用障碍(Ⅱ型)引起的高血糖症。EGCG3"Me可治疗Ⅱ型糖尿病。研究表明[24],粗多酚、EGCG和EGCG3"Me对α-葡萄糖苷酶有很强的抑制作用,半抑制浓度(IC50)依次为0.178、0.040、0.064 mg/mL,抑制类型是可逆抑制,但粗多酚和EGCG是非竞争抑制,EGCG3"Me为竞争性抑制,有更强的抑制效果。
2.4 防肥胖
EGCG3"Me对消化酶具有抑制和猝灭作用[25],可防肥胖。研究表明[22,26],EGCG3"Me抑制小鼠前脂肪细胞3T3-L1增殖和分化,影响氨基酸合成、双组分体系、ABC转录器等代谢途径从而减轻体重,比EGCG具有较高的减肥效果和较低的细胞毒性活性。Yang Y[26]等用不同浓度的EGCG3"Me(20、40、80 μg/mL)处理小鼠前脂肪细胞3T3-L1,脂质累计量分别下降到89.42±2.66%、64.36±3.13%和39.37±2.79%。此外,EGCG3"Me纳米复合物能提高EGCG3"Me的生物活性并预防肥胖相关的代谢紊乱[23]。
2.5 降血压
血管紧张素转化酶(ACE)催化血管紧张素(十肽)水解成八肽的血管紧张素Ⅱ,导致血管收缩和血压升高。临床试验表明[7],富含EGCG3"Me的Benifuuki茶比不含EGCG3"Me的茶有更好的降血压作用,主要是EGCG3"Me能抑制ACE的活性,达到降血压的作用,且EGCG3"Me在人体内的具有良好的吸收率和稳定性。
2.6 保护肝脏
EGCG3"Me可以有效减缓由酒精引起的浓度依赖性变化,每天服用一定量的EGCG3"Me,可以降低血清中天冬氨酸转氨酶(AST)、丙氨酸转氨酶(ALT)的浓度,以及肝脏细胞中丙二醛(MAD)的含量,同时,可以恢复肝脏中超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-Px)浓度到一定水平,从而保护肝脏细胞[26]。EGCG3"Me还能有效预防肝细胞冻伤,具备抗细胞毒素作用并维持细胞活性[27]。
2.7 治哮喘
EGCG3"Me对哮喘病有一定的治疗作用。与EGCG相比,EGCG3"Me对人体内一氧化氮水平和一氧化氮合酶(i-NOS)的表达具有更强的抑制作用[17],此外,由于高浓度的一氧化氮导致的DNA破坏、支气管上皮细胞脱落和功能变性和微血管渗出增加的症状减少,达到减轻气道炎症的作用。
3 茶叶EGCG3"Me含量影响因素
目前已有研究表明,茶树品种、生长环境、成熟度、采摘时间和采摘季节、加工方式等因素均会影响茶树体内EGCG3"Me含量[28]。影响EGCG3"Me含量的因素复杂,通过大量的科研实验,其影响规律逐渐明晰,今后的实验将更加系统、全面和深入。
3.1 茶树品种
茶叶中EGCG3"Me的含量根据茶树种质的不同而差异很大,中叶种含量高,小叶种含量次之,而大叶种中含量甚微[29]。绿茶和乌龙茶种质中普遍含有。高EGCG3"Me茶树种质资源的分布具有一定的区域特征,南方茶区含EGCG3"Me的茶树种质较多[30],如广东、福建,四川、重庆种质资源中也较多含有EGCG3"Me。
李建华等[31,32]采用高效液相色谱法(HPLC)对四川70份茶树品种资源一芽二叶秋梢中EGCG3"Me含量进行分析,结果发现,蜀茶5号、金观音、城西11号、凤凰单枞、崇枇71-1富含EGCG3"Me(含量>10 mg/g)。此外,只有3份种质在8~10 mg/g之间,分别是肉桂、川农2号、蒙山11号;含量在6~8 mg/g种资源有1份,为铁观音;5份茶树种质含量在4~6 mg/g,分别为浙农117、蒙山29号、锡茶5号、黄叶早、霞浦元宵茶;18份茶树种质含量在0~4 mg/g,分别为英红1号、金萱、黄金桂等。吕海鹏等[33]采用HPLC法对国内200份高多酚茶树种质的一芽二叶新梢的EGCG3"Me含量及其变化规律进行了研究,结果发现,6份茶树种质的EGCG3"Me的含量大于1%,含量最高的为TRI-1,为1.431%,此外,研究认为“紫娟”是富含EGCG3"Me的特异茶树种质,EGCG3"Me在其鲜叶中含量为1.05%[34]。唐娜等[35]对湖南的117个茶样进行种质筛选,含量最高的是C48,含量为1.7%。
3.2 生长环境
同一时期,不同地区的生态环境对茶树EGCG3"Me含量变化有显著影响。汪毅等[29]研究发现,分别生长不同地区的同一品种,其EGCG3"Me含量相差6%左右,由此认为低海拔和高光照条件对EGCG3"Me的形成非常重要。JI等[36]用黑色聚乙烯膜对鲜叶进行不同程度(90%、95%、98%和100%)、不同时间(10、20天)的遮阴处理,研究结果表明遮阴能抑制EGCG3"Me的形成,遮阴程度对EGCG3"Me含量的影响不大。陈榛淯[37]等研究发现小绿叶蝉吸食后的茶叶EGCG3"Me含量增加。
3.3 生长部位和成熟度
普遍研究发现,随着叶片成熟度的增加,茶叶中EGCG3"Me的含量有增加趋势,一般来说,茶树新梢的第三叶或者第四叶含量达到最高水平[38],少数研究发现第二叶含量最高,茎中含量很少甚至无检出。唐娜[39]发现EGCG3"Me在一芽四叶和一芽三叶中的含量最高,其次是一芽五叶,茶梗中含量极低。吕海鹏等[34]对一芽五叶新梢不同嫩度的鲜叶进行测定,发现第三叶的EGCG3"Me含量最高。李银花[1]研究金牡丹茶树品种不同成熟度叶片中EGCG3"Me含量的变化,发现茶树中EGCG3"Me含量随成熟度的增加而增加,第四叶中达最高,第五叶略有下降。而杜鸿标[40]、李建华等[32]发现金牡丹、蜀茶5号中EGCG3"Me含量随叶片成熟度先增加,第二叶含量最高。结果不一致可能由于样品来源不一致或取样方式造成了误差。
3.4 采摘时间和季节
采摘时间影响茶叶中EGCG"Me的含量。一些研究得出结论[41],在一天内,上午9点EGCG3"Me含量开始下降,到下午6点升至最高值。也有研究表明[38],在中午12点采摘的茶鲜叶中EGCG 3"Me含量明显高于早上8点和10点。因此,中午、下午或傍晚时采摘的茶叶EGCG"Me的含量较高,具体时间有待进一步研究。
已有研究普遍表明,茶叶中EGCG3"Me含量秋季>夏季>春季。李建华等[32]比较蜀茶5号同等叶位春、夏、秋三季EGCG3"Me含量,结果显示增加的趋势,汪毅等[29]发现同一品种,秋茶的EGCG3"Me含量比春茶、夏茶高出20%左右,夏茶次之、春茶最低。李银花等[1]研究两个富含EGCG3"Me茶树种质(金牡丹和水仙)春、夏、秋三个季节茶树中EGCG3"Me含量的变化,发现秋季含量最高,其次是夏季、春季。吕海鹏等[33]研究发现秋季新梢中EGCG3"Me的含量高于夏季新梢。孙业良等[38]研究发现,秋冬季茶鲜叶中的EGCG3"Me的含量高于春夏季。但也有研究结果不一致的情况,如唐娜等[39]的研究认为夏季含量相对最高,秋季次之,春季最低,结果不一致可能与其样品来源不一致或取样方式等有关。
3.5 生产工艺
加工工艺对茶叶EGCG3"Me含量有显著影响,在不同加工工艺下,EGCG3"Me含量从高到低为白茶、绿茶、黄茶、青茶、红茶。郭子豪等[42]以五种台湾特色茶树种质(青心乌龙、台茶12号、大叶乌龙、香椽种、及永康山野生茶)进行四种不同加工(绿茶、包种茶、红茶、佳叶龙茶),研究发现不同加工方式的确会造成EGCG3"Me与主要儿茶素含量的不同。罗正飞等[43]发现采用相同原料加工成绿茶、白茶以及青茶,其EGCG3"Me的含量差异不大,在加工过程中可保持相对稳定,而加工成的红茶和普洱茶中则不含有EGCG3"Me。汪毅等[29]也用同一品种制成不同茶类,发现绿茶中EGCG3"Me含量最高,其次是青茶,仅为绿茶中EGCG3"Me含量的一半左右,在红茶中很难发现EGCG3"Me。李建华等[44]用高效液相色谱-二极管阵列法检测四川绿茶、黄茶、黑茶、红茶和青茶成品茶,发现青茶EGCG3"Me含量最高,在3.14~6.08 mg/g之间,平均值为4.40 mg/g;黑茶EGCG3"Me含量在0~1.72 mg/g,平均值为0.35 mg/g;绿茶和黄茶中EGCG3"Me含量较低;绿茶中含量在0~0.39 mg/g之间,平均值为0.10 mg/g;黄茶中含量在0~0.09 mg/g之间,平均值为0.05 mg/g,红茶中未检出,造成与前人结果差异大的原因极可能是成品茶原料不一致。随后,其又以蜀茶5号秋梢加工成4种茶类,发现EGCG3"Me在绿茶、黄茶中含量最高,与鲜叶相当,其次为青茶,而红茶中未检出[32]。
不同的加工工序影响茶叶中EGCG3"Me含量。研究认为[39],不同加工工序对绿茶加工过程中茶叶EGCG3"Me的含量影响极显著,摊放叶时EGCG3"Me含量最高,杀青叶时有所下降,揉捻叶时基本不变,干燥叶时又小幅下降。萎凋工序中茶叶中EGCG3"Me含量有明显增加[32,33],且日光萎凋的效果比室内萎凋的效果要好[30]。自然萎调情况下,随着萎凋时间的增加,EGCG3"Me的含量增加,当萎凋时间达8小时,EGCG3"Me的含量达到最大值,此后萎凋时间越长,EGCG3"Me的含量越低,当到达18小时,其含量已低于萎凋前一半[43]。摇青过程会使乌龙茶EGCG3"Me含量降低[30]。杀青工序使茶叶中EGCG3"Me含量降低。吕海鹏等[33]研究发现采用不同的杀青方式加工绿茶,EGCG3"Me含量由高到低分别为蒸青绿茶、晒青绿茶、炒青绿茶、烘青绿茶,蒸青、晒青能有效保留EGCG3"Me。绿茶、黄茶、青茶揉捻工序可显著提高EGCG3"Me含量,这是由于加工过程中茶叶细胞膜被破坏,细胞内的甲基化EGCG更易被提取出来[17],而红茶揉捻工序EGCG3"Me含量显著下降,其原因一是以酶促氧化的发酵工序促使了EGCG3"Me甲基被取代或降解[43],二是红茶揉捻时发酵就已开始,研究表明[30],发酵可以减少相比于鲜叶一半的EGCG3"Me量,此时发酵破坏的EGCG3"Me量远大于揉捻增加的量[32]。干燥对茶叶中EGCG3"Me含量也有很大影响。鲜叶在不同温度(60、90、120 ℃)直接烘干,90 ℃和120 ℃干燥的茶样都明显高于60 ℃干燥的茶样[38]。吕海鹏等[34]研究发现,90 ℃的烘干温度最有利于保持茶鲜叶中的EGCG 3"Me含量。
4 EGCG3"Me合成路径探索
4.1 前体物质
根据相关体外合成实验猜测,EGCG3"Me是EGCG合成后3"位甲基化或者是合成前没食子酰基甲基化或其它前体甲基化。研究表明,茶叶中EGCG3"Me可以通过EGCG在某种甲基化酶的催化作用下转化。伍妍俊[45]实验发现,在人工模拟胃液中,EGCG3"Me的同分异构体EGCG4"Me脱去甲基,形成EGCG。苑建[46]通过合成中间体甲基化EGC,从而再合成甲基化EGCG。
4.2 酶调控
甲基转移酶调控生物体许多重要生理环节,如抑制或关闭基因表达,修复DNA损伤等,其通常以S-腺苷-甲硫氨酸(SAM)作为甲基供体催化甲基化反应,并且根据底物的作用位点分为不同类型的酶,其中,O-甲基转移酶是植物中非常重要的酶,影响植物的生长发育并调节植物与环境的相互作用,其作用底物主要是酚类物质,将SAM的甲基转移到底物的酚羟基上形成甲基化产物[47]。
吕海鹏等[33]认为,绿茶和青茶中的EGCG3"Me含量高可能与茶叶鲜叶中的甲基化酶的活性有关,而红茶和普洱茶的EGCG3"Me含量低可能与茶叶发酵(或后发酵)过程EGCG3"Me发生氧化聚合或者产生了某些去甲基化酶使EGCG3"Me向EGCG转化有关。有研究认为[48],茶树咖啡酰辅酶A氧甲基转移酶(CsCCoAOMT)催化甲基化EGCG合成。李银花[1]研究发现基因CsCOMT的表达水平与茶树中EGCG3"Me的含量呈正相关,通过SMART RACE技术成功克隆了甲基化EGCG合成酶的全长cDNA序列,构建了该基因的原核表达载体。费冬梅[14]首次对产EGCG-O-甲基转移酶的重组大肠杆菌的培养条件研究,得到了最佳培养条件。
4.3 受热变化
高温等因素引起的差向异构化也可能是茶叶加工过程中EGCG3"Me含量降低的原因之一。长时间高温处理以及提取和灭菌中的热处理等过程中,EGCG3"Me差向异构化为GCG3"Me,目前在溶液中已经证实了这种转化的存在。当茶叶发酵时可能导致EGCG3"Me被进一步氧化聚合,部分生成甲基化的茶黄素[33,34,49]。
4.4 关联代谢途径
茶叶EGCG3"Me代谢合成途径的研究多集中在儿茶素代谢途径。杜鸿标等[40]研究了金观音、金牡丹、黄玫瑰三个品种不同季节、不同发育阶段中儿茶素合成代谢途径的相关基因CHS、CHI、PAL、C4H、ANS等与EGCG3"Me的表达趋势符合情况,不同品种基因表达量变化趋势不一,相关性不强。这可能因为富含EGCG3"Me的品种中个别儿茶素化合物的代谢不同于其他茶树品种[36,50]。Luo Y等[51]鉴定到了一个bHLH转录因子差异表达基因CsbHLH62,其能够抑制与EGCG3"Me的积累呈正相关的CCoAOMT、CsLAR和CsDFR的表达进而负向调控EGCG3"Me的合成。也有研究表明[52],EGCG3"Me的含量和咖啡碱代谢无相关关系。
5 展望
EGCG3"Me具有不可替代的抗过敏等保健功效,具有进行深加工、开发为保健产品的潜力和价值,但茶树体内天然EGCG3"Me含量低,化学合成多用危害、剧毒的试剂难于处理副产物,酶法合成绿色、健康但效率低且不可控,今后对EGCG3"Me分离、合成的研究将集中在绿色、高效、专一产物方法的开发。高EGCG3"Me含量茶树资源调查较少,急需挖掘高EGCG3"Me的茶树种质资源为EGCG3"Me实验和深加工提供材料,为种质资源的开发提供借鉴依据。EGCG3"Me含量的影响因素有很多,但具体的影响机理尚不清楚,甲基化EGCG代谢机理的研究主要集中在甲基转移酶,大多为体外合成实验,针对甲基化EGCG的体内实验较少,茶树体内EGCG3"Me代谢合成机理尚不清楚,同位素标记示踪实验、茶树体外分解实验也将展开,随着组学研究热潮的升温,组学也将应用于EGCG3"Me的研究。随后EGCG3"Me深加工保健产品也将逐渐被开发,广泛推向市场,解决困扰人们的过敏问题。
