低出生重对仔猪肝脏抗氧化功能的影响及精氨酸的营养效应
2019-07-24田一航陈代文毛湘冰罗玉衡黄志清罗钧秋
田一航,宋 毅,陈代文,余 冰,何 军,虞 洁,毛湘冰,罗玉衡,黄志清,罗钧秋,郑 萍
(四川农业大学动物营养研究所/动物抗病营养教育部重点实验室,四川 雅安 625014)
遗传育种技术的发展和饲养管理的改善虽然提高了母猪的窝产仔数,但是也提高了低出生重(low birth weigh,LBW) 仔猪数量[1],自然发生的LBW 仔猪比例高达 15%~20%[2-3]。LBW 显著提高仔猪断奶前死亡率和发育不良比例[4],造成饲养管理所投入的精力和资本增加。宫内发育迟缓(intrauterine growth retardation,IUGR) 即哺乳动物妊娠期胚胎或胎儿发育受阻,是LBW 仔猪产生的主要原因[2,5]。仔猪出生前,生长于氧气浓度相对较低的子宫,出生后,则会吸入高氧气浓度的空气,此转变会引起其氧化还原失衡[6-7],随着其抗氧化系统的发育,氧化还原平衡可逐渐恢复[8]。LBW 仔猪抗氧化系统的发育可能异常,表现为抗氧化能力低于NBW 仔猪。与NBW 仔猪相比,IUGR 新生仔猪抗氧化能力显著降低[9-10],IUGR 哺乳仔猪肝脏GPx 活性显著降低,MDA 含量显著提高[11],断奶两周的LBW仔猪血浆Fe3+还原能力和GPx 活性显著降低[5]。
Arg 是幼龄哺乳动物所必需的氨基酸,在体内既参与蛋白质的合成,又是NO、多胺、肌酸和胍丁胺合成的前体[12]。哺乳仔猪出生7日龄到14日龄期间,血液循环中Arg 及其前体瓜氨酸、鸟氨酸的含量显著降低[13-14]。前人研究表明,一头7日龄的哺乳仔猪每天Arg 需要量为2.7 g,而猪乳只能提供1.06 g[15-16]。Arg 有改善仔猪抗氧化功能的作用。研究表明,饲粮添加Arg 显著降低仔猪血清MDA 含量,缓解断奶引起的氧化应激[17]。饲粮添加Arg 可缓解Diquat 诱导的氧化损伤,增强仔猪的抗氧化能力,缓解氧化应激[18]。饲粮添加4%的Arg 显著降低血清MDA 含量,提高血清 GPx 活力,0.8%的 Arg 显著提高血清CAT、SOD 和GPx 活力,从而缓解氧化鱼油引起的氧化应激[19]。
LBW 仔猪抗氧化能力降低,而Arg 有改善仔猪抗氧化功能的作用。因此本试验通过比较LBW 仔猪和NBW 仔猪肝脏抗氧化能力,并在饲粮中添加Arg 饲喂 LBW 仔猪,考察 Arg 添加对 LBW 哺乳仔猪肝脏抗氧化功能的影响。
1 材料和方法
1.1 试验动物与试验设计
本研究包括2 个试验。所用仔猪从体况相近、胎次相近和产期一致的母猪所产DLY 新生仔猪中选择,根据前人研究确定选择标准[20-21]。试验1 选取10 对公猪,从同一窝中选到的NBW 和LBW 为1 对,两组仔猪体重分别为(1.66±0.07)和(0.83±0.04) kg,不吃初乳,2~4 h 内麻醉屠宰。试验 2 从 72 窝仔猪中标记出16 头NBW 仔猪和72 头LBW 仔猪,待其吃完初乳,4日龄时,再按照体重接近和性别比例一致的原则从标记过的仔猪中选取10 头NBW 和20头 LBW 仔猪,NBW 仔猪饲喂基础饲粮,将 20 头LBW 仔猪分为两组,分别饲喂基础饲粮和基础饲粮中添加1.0% L-Arg 的饲粮,均配制成人工乳,各组仔猪初始体重分别为(1.96±0.03)、(1.05±0.04)和(1.05±0.04) kg,饲喂 21 d,试验第22 天早晨进行麻醉屠宰。
1.2 试验材料
L-Arg 由日本味之素公司提供,纯度为99%,LAla 由上海易蒙斯公司提供,纯度为99%。
1.3 试验饲粮
饲粮为人工乳,由代乳粉加水配制而成,按照代乳粉:水=1:4 的比例加40℃温开水,充分溶解混匀后饲喂。基础饲粮参考前人[22-23]研究配制,添加1.0%L-Arg 的饲粮是在基础饲粮中添加1.0%的L-Arg,并通过添加葡萄糖和L-Ala 使两组饲粮等能等氮,此添加量根据本课题组前期研究确定,基础饲粮组成及营养水平见表1。
1.4 饲养管理
试验在四川农业大学动物营养研究所科研基地进行,所有仔猪单笼饲养于代谢笼中。试验前对圈舍及代谢笼进行全面消毒,试验第一周温度控制在31~32 ℃,之后每周降低 2 ℃,相对湿度控制在50%~60%。每天饲喂时间为 06:00,09:00,12:00,15:00,18:00,21:00,24:00,每次以仔猪吃饱为准。试验期间不对猪只使用任何抗生素类药物,其余按基地要求进行操作管理。

表1 基础饲粮组成及营养水平Table 1 Composition and nutrient levels of basal diets %
1.5 样品采集与处理
屠宰后,迅速打开仔猪腹腔,分离出肝脏,并取适量组织样装于冻存管中,锡箔纸包好,投入液氮速冻,并放入-80 ℃冰箱保存待测。
1.6 测定指标与方法
1.6.1 肝脏抗氧化能力相关指标测定
使用南京建成生物工程研究所的试剂盒,测定肝脏组织中ASA 和AHR 活性,抗氧化酶SOD、GPx活性和MDA 的含量,严格按照说明书进行操作。先取约1 g 组织样品,加入9 倍体积生理盐水,冰水浴条件下机械匀浆,2 500 r/min 离心10 min,然后对上清液总蛋白质含量进行测定,最后取上清液进行各个指标的测定,测定结果定义为,每毫克蛋白质中该指标的活性或含量。
1.6.2 基因表达测定
实时定量PCR 法测定哺乳仔猪肝脏基因,包括核因子NF-E2 相关因子(Nrf2)、血红素氧合酶1(HO-1)、醌氧化还原酶 1(NQO-1)、白细胞介素1β(IL-1β)、白细胞介素 2(IL-2)、白细胞介素 6(IL-6)、白细胞介素 10(IL-10)、肿瘤坏死因子 α(TNF-α)、干扰素γ(IFN-γ)的相对表达量。
总 RNA 的提取按照试剂盒(Trizol Reagent,TakaPa,日本)操作说明进行,RNA 质量检测使用核酸蛋白检测仪 (Beckman DU-800,CA,美国) 于 260 nm 检测,A260/A280 表示 RNA 的纯度,该比值在1.8~2.0 说明 RNA 纯度较好。cDNA 的合成按照逆转录试剂盒(Prime ScriptTMregent kit,TaKaRa,日本),反应结束后-20℃保存待用。利用美国国家生物技术信息中心(NCBI)搜索基因序列,运用Primer 5 进行引物设计,由上海生工生物工程公司合成,引物序列见表2。用实时定量PCR 仪(ABI7900HT Real-Time PCR System,ABI,美国)进行测定,反应荧光染料为SYBR Green I(TaKaRa,日本)。反应体系为 10 μL:5 μL SYBR Premix Ex TaqTMII(2×),0.5 μL 上游引物,0.5 μL 下游引物,3 μL 双蒸水,1 μL cDNA 模板。以β-actin 和18sRNA(取两者CT 值的几何平均值)为内参基因,使用2-ΔΔCT方法计算目的基因的相对表达量。
1.7 数据统计分析
试验数据首先用Excel 2010 进行初步整理,然后使用SPSS 21.0 软件进行数据分析,新生仔猪采用配对样本t 检验,使用P 表示其显著性。哺乳仔猪采用独立样本t 检验,两两比较,LBW 哺乳仔猪与NBW 哺乳仔猪比较的显著性用P1 表示,饲粮添加1.0% Arg 饲喂的哺乳仔猪与LBW 哺乳仔猪比较的显著性用 P2 表示。以 P<0.05 为差异显著,0.05≤P<0.1 为有趋势。结果以“平均值±标准误”表示。

表2 实时定量PCR 引物序列及参数Table 2 Sequences and parameters of primers for the real-time qPCR
2 结果与分析
2.1 新生仔猪的体重
从表3可以看出,与NBW 新生仔猪相比,LBW新生仔体重显著降低(P<0.05)。

表3 新生仔猪的体重Table 3 Body weight of neonatal piglets
2.2 低出生重对新生仔猪肝脏抗氧化能力的影响
从表4可以看出,与NBW 新生仔猪相比,LBW新生仔猪 T-SOD 和 GSH-Px 活性显著降低(P<0.05),AHR 活性有降低的趋势(P=0.08)。

表4 低出生重对新生仔猪肝脏抗氧化能力的影响Table 4 Effects of LBW on the antioxidant activity in liver of neonatal piglets
2.3 低出生重对哺乳仔猪肝脏抗氧化功能的影响及精氨酸的营养效应
从表5可以看出,与NBW 哺乳仔猪相比,LBW哺乳仔猪肝脏 ASA 和 AHR 活性显著降低(P<0.05);与LBW 哺乳仔猪相比,饲粮添加1.0%Arg 显著提高 LBW 哺乳仔猪肝脏 AHR、T-SOD 和 GSH-Px 活性(P<0.05)。
2.4 低出生重对哺乳仔猪肝脏抗氧化相关基因表达的影响及精氨酸的营养效应
从表6可以看出,与NBW 哺乳仔猪相比,LBW哺乳仔猪肝脏Nrf2 和NQO-1 表达量显著提高(P<0.05);与LBW 哺乳仔猪相比,饲粮添加1.0%Arg显著降低 LBW 哺乳仔猪肝脏 NQO-1 表达量(P<0.05)。

表5 低出生重对哺乳仔猪肝脏抗氧化功能的影响及精氨酸的营养效应Table 5 Effects of LBW on the antioxidant activity in liver of suckling piglets and the nutritional regulation by L-arginine

表6 低出生重对哺乳仔猪肝脏抗氧化相关基因的影响及精氨酸的营养效应Table 6 Effects of LBW on the expression of antioxidant genes in liver of suckling piglets and the nutritional regulation by L-arginine
2.5 低出生重对哺乳仔猪肝脏细胞因子基因表达的影响及精氨酸的营养效应
从表7可以看出,与NBW 哺乳仔猪相比,LBW哺乳仔猪肝脏 IL-1β(P=0.09)和 IL-6(P=0.06)表达量有提高的趋势;与LBW 哺乳仔猪相比相比,饲粮添加1.0%Arg 显著降低LBW 哺乳仔猪肝脏IL-1β和 IL-6 表达量(P<0.05)。
3 讨论与结论
3.1 低出生重对新生仔猪肝脏抗氧化功能的影响
自由基产生于机体代谢,过多的自由基会导致脂质过氧化、蛋白质变性和DNA 损伤[24]。ASA 和AHR 活性分别代表超氧阴离子和羟自由基的清除能力[25]。线粒体电子传递过程产生的自由基是机体自由基的主要来源,肝脏富含线粒体,是自由基攻击的主要器官[26-27]。在生理条件下,机体抗氧化系统可清除自由基,维持氧化还原平衡。前人运用蛋白组学研究表明,与NBW 新生仔猪相比,IUGR 新生仔猪肝脏抗氧化相关蛋白含量显著降低[9]。有研究结果表明,与NBW 新生仔猪相比,IUGR 新生仔猪肝脏抗氧化酶T-SOD、GPx 和CAT 活性显著降低[10]。本研究结果表明,与 NBW 相比,LBW 新生仔猪肝脏T-SOD 和 GSH-Px 活性显著降低,AHR 活性有降低的趋势,本研究结果与前人一致,说明LBW新生仔猪肝脏抗氧化能力降低。
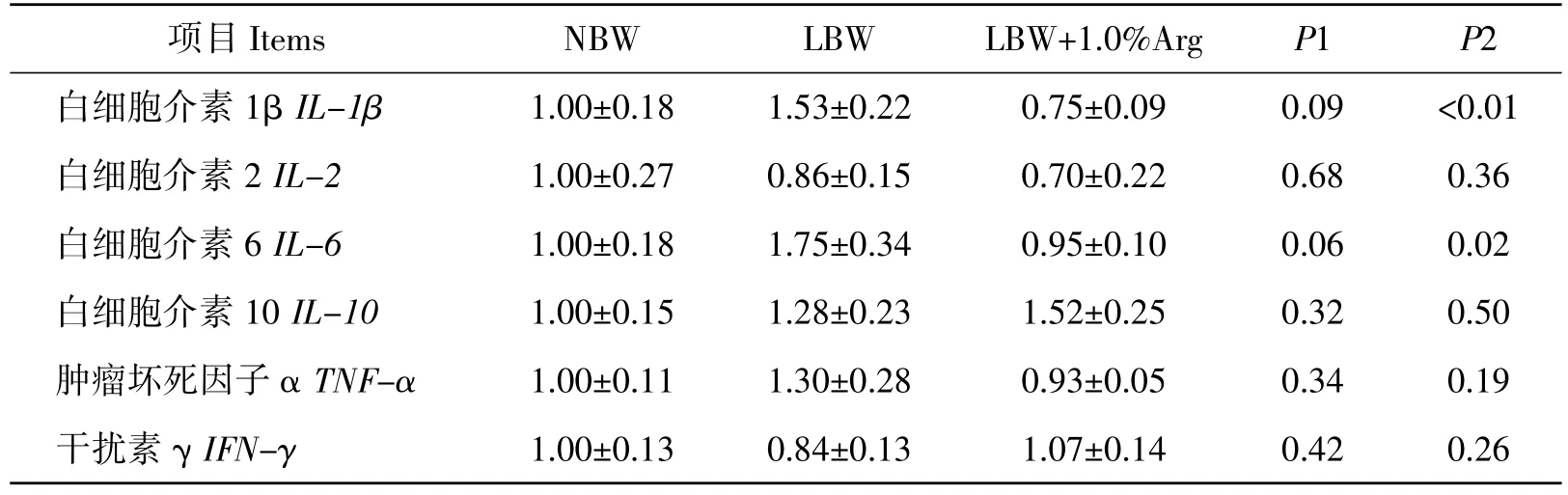
表7 低出生重对哺乳仔猪肝脏细胞因子基因表达的影响及精氨酸的营养效应Table 7 Effects of LBW on the expression of cytokine genes in liver of suckling piglets and the nutritional regulation by L-arginine
3.2 低出生重对哺乳仔猪肝脏抗氧化功能的影响及精氨酸的营养效应
随着日龄的增加,LBW 仔猪肝脏抗氧化能力可能仍然低于NBW 仔猪。前人研究结果表明[28],与NBW 哺乳仔猪相比,IUGR 哺乳仔猪7日龄时肝脏总抗氧化活性和CAT 活性显著降低,23日龄时肝脏总抗氧化活性和SOD 活性显著降低。有研究结果表明,与NBW 哺乳仔猪相比,IUGR 哺乳仔猪28日龄时肝脏GPx 活力显著降低[11,29]。本研究结果表明,4日龄开始,饲喂 21 d 后,与 NBW 仔猪相比,LBW仔猪ASA 和AHR 活性显著降低,本研究结果与前人一致,说明LBW 哺乳仔猪肝脏抗氧化能力降低。我们前期研究表明[30],4日龄开始,人工乳饲喂21 d后,与NBW 仔猪哺乳相比,LBW 哺乳仔猪末重、平均日增重、平均日采食量和料重比显著降低,而饲粮添加1.0%Arg 显著提高LBW 哺乳仔猪末重、平均日增重和平均日采食量,有提高其料重比的趋势,说明LBW 哺乳仔猪生长性能降低,补充1.0%Arg 可改善其生长性能。Arg 有改善仔猪抗氧化功能的作用。前人研究表明,Arg 添加提高氧化应激仔猪血清总抗氧化活性、GPx 活性和 SOD 活性[18]。前人研究表明,添加0.8%的Arg 显著提高了血清CAT、SOD 和 GPx 的活性,提高仔猪抗氧化能力[31]。本研究结果表明,与LBW 组仔猪相比,饲粮添加1.0%Arg 显著提高LBW 哺乳仔猪肝脏T-SOD、GSH-Px和AHR 活性,本研究结果与前人一致,说明补充1.0%Arg 改善了LBW 哺乳仔猪肝脏抗氧化功能。
Nrf2 是机体发挥抗氧化作用重要的转录因子,氧自由基的大量产生使Nrf2 活化,活化的Nrf2 进入细胞核与DNA 启动子区域的抗氧化元件结合,激活下游的Ⅱ相解毒酶和抗氧化酶的转录,通过调节NQO1、HO-1、SOD 等基因的转录,可抵抗自由基造成的危害[32-33]。NQO-1 在细胞内属于高度可诱导的还原酶,主要受Nrf2 信号途径的调节,它以NADH 或NADPH 为电子供体,催化醌类化合物发生还原反应,从而阻止它们进一步参与氧化还原反应和氧自由基的产生[34]。仔猪受到应激后,启动自身的保护性反应,显著提高肝脏Nrf2 蛋白表达量[35]。本研究结果表明,与NBW 哺乳仔猪相比,LBW 哺乳仔猪肝脏Nrf2 和NQO-1 表达量显著提高,与LBW 哺乳仔猪相比,饲粮添加1.0%Arg 显著降低LBW 哺乳仔猪肝脏NQO-1 表达量,说明LBW 哺乳仔猪启动了自身的保护性反应,激活了肝脏Nrf2 途径,补充1.0%Arg 可能缓解了此途径的激活,改善LBW 哺乳仔猪肝脏抗氧化功能。
3.3 低出生重对哺乳仔猪肝脏细胞因子基因表达的影响及精氨酸的营养效应
细胞因子主要是由免疫细胞(如单核、巨噬细胞、T 细胞、B 细胞、NK 细胞等)经刺激而合成、分泌的一类具有广泛生物学活性的小分子蛋白质,其中IL-1β、IL-2、IL-6、TNF-α 和 IFN-γ 属于炎性细胞因子,可引发炎症,IL-10 属于抗炎细胞因子,可抵抗炎症。Zheng P.等研究表明,与NBW 仔猪相比,IUGR 仔猪肝脏 IL-1β 和 TNF-α 含量显著提高,IL-10 含量显著降低导致肝脏发生炎症[28],本研究结果与文献相似,与NBW 组相比,LBW 组哺乳仔猪肝脏IL-1β 和IL-6 表达量有提高的趋势,说明LBW 哺乳仔猪肝脏可能发生了炎症。饲粮添加Arg可调节仔猪细胞因子表达。Li Q.等研究结果表明,饲粮添加Arg 可通过TLR4-NF-ΚB 信号途径降低仔猪肝脏促炎因子的分泌,缓解大肠杆菌诱导的肝脏损伤[36]。本研究结果表明,与LBW 组仔猪相比,饲粮添加1.0%Arg 显著降低LBW 哺乳仔猪肝脏IL-1β和IL-6 表达量,该结果与前人一致,说明补充1.0%Arg降低LBW 仔猪肝脏炎性细胞因子的表达,有保护肝脏的作用。
综上,在本试验条件下,LBW 仔猪肝脏抗氧化能力低于NBW 仔猪,补充1.0% Arg 可改善LBW哺乳仔猪肝脏抗氧化功能。
