被子植物MADS-box基因研究进展
2019-07-24王鹏洋付泽元曲姗姗
王鹏洋,付泽元,曲姗姗,梁 源
(东北林业大学 生命科学学院,黑龙江哈尔滨 150040)
0 引言
MADS-box基因作为重要的调控基因,在植物生长和发育的许多方面发挥着重要的调节功能[1]。现如今,已经在许多被子植物中发现数以百计的MADS-box基因[2]。MADS-box蛋白作为一种转录调控因子,在控制植物开花时间、花器官形成和分生组织特异性等方面的信号转导过程中起着关键作用[3]。MADS-box基因还是一类家族基因的总称,该家族基因在长期进化的过程中发生了不同程度的重复事件,这就导致了MADS-box基因家族成员具有功能差异,即亚功能化或新功能化[4]。MADS-box基因的名称是由4个物种的重要转录因子名字的第1位字母拼接而成,它们分别是:M来源于酵母的MCM1,A为调控拟南芥花器官形成的AG,D是金鱼草的DEF,S是人血清应答因子SRF[5]。在被子植物中MADS-box是一个巨大的基因家族,该家族成员分散在被子植物整个基因组中。
1 MADS-box基因的分布
MADS-box基因分布十分广泛,在植物、动物、微生物中都已经有学者将其克隆出来。在植物中,MADS-box基因遍布整个植物界。在被子植物(如水稻、小麦、玉米等) 单子叶植物和核心双子叶植物拟南芥中都有大量的分布[6]。在植物生长发育的不同时期,均有MADS-box家族的基因表达。在植物不同的部位(如根、茎、叶) 或者作为生殖器官的花、果、种子中,不同的MADS-box基因会有不同的特异性表达[7],以实现特异性的调控作用。
2 MADS-box蛋白的结构
植物MADS-box基因主要为MIKC型基因,典型的MIKC型MADS-box蛋白有相对保守的结构域,其由 MADS-box(M)、Intervening(I)、K-box(K) 和C-terminal(C) 共 4个部分组成[8]。其中,MADS-box的结构域最为保守,由60多个氨基酸残基组成,MADS-box具有与特定DNA结合的活性,同时也在二聚体和辅助因子结合中发挥作用;I区约由35个氨基酸残基构成,可变性较大,为弱保守结构域,是调控DNA结合二聚体形成的决定性因素;K-box结构域可以形成参与蛋白二聚化的两性螺旋,介导蛋白质与蛋白质的相互作用;C区位于K-box的下游末端,是最易变的区域,它参与转录激活和多转录因子复合物的形成[9]。根据I区和K-box的不同,MIKC型的MADS-box蛋白可以进一步细分为2类,分别是MIKCC型蛋白和MIKC*型蛋白[10]。MIKCC型蛋白的I结构域其基因仅有一个外显子;而MIKCC型蛋白的I结构域的基因有4~5个外显子[11]。
3 MADS-box基因的进化
研究表明,MADS-box基因在早先年代只有一个共同的祖先,而MADS-box基因家族中的基因复制被认为是进化的关键过程[12]。在基因复制之后,一个基因可以有几种不同的命运。如果一个基因是完全复制的,这通常会导致功能冗余。另一方面,一个复制的基因可以保留祖先的功能,而另一个获得突变成为伪基因,在这种情况下如果获得突变的基因是有益的积极突变,就会被自然所选择,从而产生稳定的新功能;还有一种可能性是,这2个基因都获得了功能缺失突变,然而突变结果是2个基因功能互补,2个基因会在长期进化的过程中一同保留下来,因为它们在一起才能实现它们唯一祖先的原始功能,这也被称为复制-退化-互补 (DDC) 模型[13]。以上这些获得性突变变化分别称为去功能化、新功能化和亚功能化。不同物种间MADS-box基因家族的大多数差异也被认为是由基因复制引起的。
4 被子植物MADS-box基因的生物学功能——ABCDE模型
MADS-box基因在被子植物中是一类有着重要功能的转录调控因子,尤其在被子植物的花中至关重要,科学家对MADS-box基因的功能了解得还不够多,现在对其研究最完备的功能依然为控制植物花器官的识别和发育,尤其是在被子植物中雌雄同体的物种中。MADS-box基因通过在花的不同部位、不同时间特异性表达,控制着开花的时间、花的性别的确立、花器官的形成等重要生理过程。
在雌雄同体被子植物中,花器官的特性是由MADS-box基因编码的转录调控因子控制的,该基因在此作为花器官识别基因,被分为A,B,C,D,E共5类[14],学者对控制花器官识别遗传机制的详细分析提出了ABCDE模型。在该模型中,在花分生组织中表达不同类别的基因来决定未来形成非生殖器官的萼片(sepals) 或花瓣(petals),还是形成作为生殖器官的雄蕊 (stamens) 或心皮(carpel)。在模式植物拟南芥和其他雌雄同体物种中,共分为4轮,第1轮A类和E类基因可以一起调控萼片识别;第2轮A类结合B类和E类基因一起调控花瓣识别;第3轮B类结合C类和E类基因可明确识别雄蕊;第4轮C类与D类、E类基因可以明确识别心皮[15-16]。其中,E类基因在每一轮中都有表达,A类和C类基因的表达产物相互抑制不能同时共存,A,B,C类基因是该模型中最核心的部分。A类基因主控萼片和花瓣的识别,当A类基因失去功能时,失去对C类的抑制效应,则C类基因会一反常态的表达于第1轮与第 2轮中,第1轮的花器官识别不再是萼片而是心皮,第2轮的花器官识别会由花瓣转化为雄蕊。B类基因主要在花瓣和雄蕊的识别期发挥作用,如果B类基因失活,第2轮和第3轮就会完全由A和C类控制形成萼片和心皮。C类基因主控雄蕊和心皮的发育,如果C类基因失去功能无法抑制A类基因在此时表达,第3轮和第4轮就会生成萼片和花瓣[17-19]。
ABCDE模型见图1。
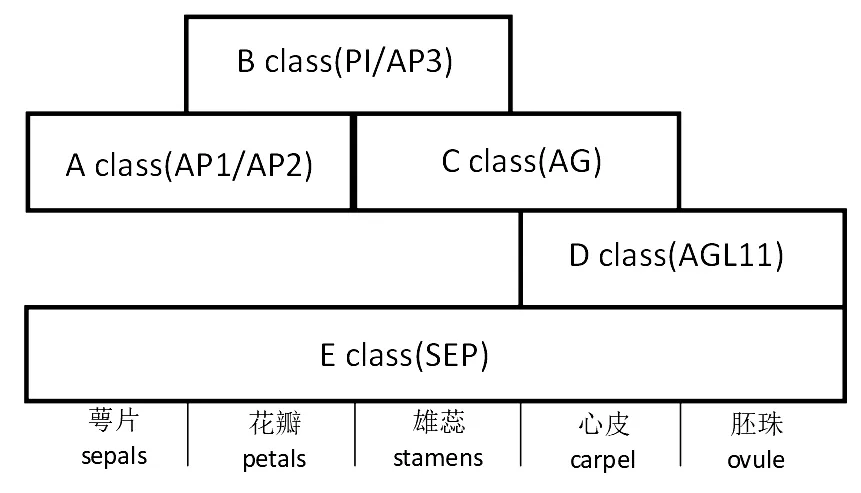
图1 ABCDE模型
MADS-box蛋白能与DNA结合的能力是其发挥功能的基础,以此决定花器官的识别,而蛋白质-蛋白质相互作用的能力也可能是花器官形成过程中可能涉及的发育过程的一部分[20]。MADS-box蛋白早先被认为是以二聚体形式存在来调控靶基因的表达。如通过酵母双杂交试验得到,PI能够与AP3或TM6结合形成异源二聚体;AG,AP1,AP3和AGL1还可以自二聚。随着进一步研究有学者提出了四聚体模型,即2组二聚体通过蛋白互作效应来组成一个四聚体结构,如在拟南芥中,AtAP1-AtAP1同源二聚体与AtSEP-AtSEP异源二聚体形成四聚体控制萼片的发育;2个异源二聚体AtAP3-AtPI和At-SEP-AtAP1形成四聚体,决定花瓣的识别;2个异源二聚体AtAP3-AtPI和AtSEP-AtAG形成的四聚体决定雄蕊的形成;AtAG-AtAG和AtSEP-AtSEP的四聚体控制心皮的发育。ABCDE模型已被成功地用于解释被子植物中花器官组织的识别,这似乎是一种较为保守的机制。
5 结语
随着生物技术的发展,目前关于MADS-box基因与花器官识别取得了重要的研究进展。因为基因重复导致MIKC型MADS-box基因的进化,进而导致这一类基因去功能化、亚功能化或新功能化。对MADS-box基因研究最多的是在花器官识别方面,在近几十年内,对被子植物中雌雄同株的物种进行了研究,指出MADS-box的ABCDE类基因是控制雄雌器官发育的分子开关之一;而在雌雄异株物种中,ABCDE基因能调控单性花的形成过程。随着研究的深入,由ABCDE引申出的滑动边界模型和模糊边界模型还可以解释裸子植物、单子叶植物等的花发育过程,这2个模型都表明,这些物种的非常规花的表型主要是由于A类和C类基因表达边界的改变而实现的。
研究MADS-box基因发挥功能的分子机制,对于解释植物花发育过程中各种生理生化现象具有重要的研究意义,在未来,可以根据MADS-box基因在花器官发育的时间、空间上特异性表达的特点来对花卉进行人工改造;对于有关于花器官的经济作物可以大量生产等应用,将理论研究置于实际应用中,创造更高的价值。
