基于网孔分解袋法的荒漠草原柠条枯落物分解过程中的养分变化
2019-07-20刘佳楠常海涛张安宁刘任涛
刘佳楠,赵 娟,常海涛,张安宁,陈 蔚,刘任涛
(宁夏大学西北退化生态系统恢复与重建教育部重点实验室,宁夏 银川 750021)
枯落物作为养分的一种基本载体,是陆地生态系统的重要组成部分与土壤物质转化的基础[1-2]。枯落物分解过程中的养分释放,在维持土壤肥力、保证植物生长繁殖所需养分、促进陆地生态系统正常的物质循环和养分平衡方面起着重要的作用[3]。柠条(Caragana korshinskii)属于豆科锦鸡儿属,落叶灌木,萌蘖力和再生能力极强,耐旱耐寒[4],不但可以改善土壤物理性质,而且能防风固沙、拦泥蓄水、减少地表径流以及维护良好的环境效益[5],是西北干旱半干旱区重要的护土植物之一。柠条灌丛枯落物的输入可以促进土壤有机质和营养物质的富集,这些营养丰富的土壤被称为“生育岛”或“资源岛”,进而在荒漠生态系统的动力学中扮演着重要角色[6]。并且,作为地上-地下生物养分循环的关键“纽带”,柠条枯落物分解对沙地、草地土壤理化性质的演变过程存在重要的影响,对于维持陆地生态系统元素平衡有着重要的意义,是陆地生态系统物质循环的重要环节[7]。因此,研究荒漠草原区柠条灌丛枯落物分解过程中养分的变化特征,对理解灌丛“肥岛”效应、人工林管理以及维持荒漠草原生态系统元素平衡具有重要意义[8]。
目前,关于不同林龄或不同发育阶段林地枯落物分解过程中元素变化动态的研究主要集中在森林生态系统[9-10]。对黄土高原地区刺槐(Robinia pseudoacacia)、柠条人工林枯落物的研究发现,不同林龄间N、P含量均表现为成熟林 > 中林龄 > 幼林龄[11]。油茶(Camellia oleifera)枯落物分解过程中N、P、K元素含量均表现为成林 > 幼林,并且季节变化差异明显[12]。分析发现,不同林龄对枯落物营养元素释放影响十分深刻,成林具有较高的营养元素释放率,有利于土壤生态系统物质循环过程[13]。不仅如此,利用网孔分解袋法限定土壤生物类群的分解作用,亦表现出不同生物类群对枯落物分解过程中营养元素变化具有重要作用,而且随分解时间呈现出显著变化。李海涛等[14]利用网袋法对亚热带地区4种人工林凋落物分解动态及养分释放进行研究,结果发现C元素在各林分中始终表现为净释放,而N元素在分解过程中伴随着释放、富集两种状态。对川滇高山栎(Quercus semecarpifolia)凋落物分解及养分释放特征的研究发现,枯落物在不同网孔年际之间分解速率存在显著差异,N、P元素出现富集→释放的变化规律,而木质素和纤维素表现为持续下降趋势[15]。但是,关于荒漠草原不同发育阶段柠条灌丛微生境枯落物分解过程中营养元素变化动态研究,报道较少,并且基于网孔分解袋法的荒漠草原枯落物分解的营养元素变化规律,尚不清楚。
柠条灌丛不同发育阶段(极小型、小型、中型和大型),其灌丛下微生境间存在显著差异性,间接影响枯落物的分解过程和营养元素释放[2],并且不同微生境的差异性亦影响到参与枯落物分解过程的土壤动物类群分布,进而影响到灌丛微生境中枯落物分解与养分循环[6]。鉴于此,本研究选择宁夏荒漠草原区不同发育阶段柠条枯落物为研究对象,采用不同网孔分解袋法研究不同发育阶段柠条灌丛枯落物营养元素的变化规律,旨在揭示荒漠草原柠条灌丛枯落物分解过程中元素的释放与富集对土壤养分循环的影响,为荒漠草原生态系统生物地球化学循环过程解析、人工林建设与沙漠化防治提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于宁夏回族自治区盐池县,该区地处内蒙古自治区、甘肃省和陕西省的交界处,位于毛乌素沙地南缘,地理坐标为 106°59′-107°07′ E,37°56′-38°02′ N,平均海拔 1 400 m。该区域属典型温带大陆性季风气候区,年平均气温7.89 ℃。年降水量250~350 mm。年均无霜期为165 d,年平均风速为 4.89 m·s-1。
研究样地为该县域东北方向10 km处(107°27′ E,37°49′ N,海拔 1 416 m)。土壤表面为风沙土覆盖,土壤粗砂粒含量超过90%,土壤肥力较低。主要植物有柠条、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)、赖草 (Leymus secalinus)、胡枝子(Lespedeza bicolor)、猪毛蒿(Artemisia scoparia)、牛枝子 (Lespedeza potaninii)和沙蓬 (Agriophyllum squarrosum)等。
1.2 研究方法
1.2.1 试验设计与分解袋布设
于2016年10月,在研究样地内选取柠条锦鸡儿进行标记,然后根据柠条灌丛冠幅大小(高度 ×冠幅),选取生长健硕的灌丛植株进行标记,分别用Ⅰ、Ⅱ、Ⅲ、Ⅳ表示极小型(71 cm × 115 cm)、小型 (122 cm × 189 cm)、中型 (155 cm × 245 cm)、大型(223 cm × 341 cm)柠条灌丛,以代表不同大小柠条锦鸡儿灌丛微生境演变过程。每类灌丛包括3个重复(即3株生长健旺、高度相近的样株),共有12株标记灌丛样株(3个重复 × 4种类型)。
首先,在标记样株下利用样方法收集地表枯落物,并依据地表柠条枯落物分布特征,计算出单位面积柠条枯落物干物质量为5.8 g·cm-2。然后,在相同生境中收集新鲜柠条叶片(带叶柄),带回实验室自然风干后,依据分解袋规格(15 cm × 15 cm)称取6 g分别装入4 mm(允许所有土壤动物进入)、2 mm(允许中小型土壤动物进入)和0.01 mm(无土壤动物进入)的尼龙网袋中。在分解袋上标好编码,并做好记录。枯落物袋由两层尼龙网缝合而成,贴地面层均采用0.01 mm孔径的尼龙网,避免枯落物损失和地下土壤动物的干扰。
在极小型、小型、中型、大型4种灌丛生境中布设上述3种规格的分解袋。从灌丛下正东方向开始布设,分解袋距离灌丛根部15 cm,且2个分解袋间间隔10 cm,然后,用铁丝弯勾固定,使分解袋与土壤紧密接触,且埋藏深度为0~2 cm。在2016年12月初完成分解袋布设,并于生长210 d时(2017年7月)取回枯落物分解袋36袋,即3种规格 × 3 个重复 × 4 种类型灌丛 = 36 个。
1.2.2 枯落物取样与分析
取样时,按照相同的顺序从每个灌丛下取回样品3袋,即取回36袋样品。将取回的样品装入信封,保存在恒温箱内,带回实验室。首先清除枯落物表面附着的泥沙和其他杂质,然后将取回的枯落物从分解网袋中转移到干净的信封中,置于烘箱,于70 ℃烘干48 h,称重,并记录枯落物样品的剩余干重。
将剩余枯落物样品进行粉碎,用于测定枯落物N、P、K、木质素和纤维素含量。用凯氏定氮法测定N含量[16]。用氢氧化钾熔融-钼锑抗比色法测定P含量[16]。用氢氧化钠熔融-火焰光度计法测定K含量[16]。用GBT 20805-2006范氏法测定木质素含量[17]。用NYT 1459-2007规定的方法测定纤维素含量[18]。
1.2.3 数据处理与统计分析
枯落物养分残留率(NR)计算公式为:
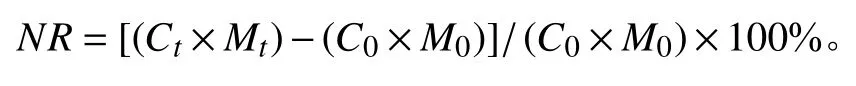
式中:Ct表示分解时间t时元素养分含量(%),Mt表示分解t时的残留干物质质量,C0表示初始元素养分含量(%),M0表示初始干物质质量[19]。
采用SPSS 20.0软件对所测数据进行统计分析,用平均值和标准误表示测定结果分别对同一网孔内不同灌丛大小、同一类型灌丛下不同网孔间养分残留率进行单因素方差分析(One-way ANOVA)和相关性分析(Pearson),作图用Excel 2016完成。
2 结果与分析
2.1 枯落物初始质量特征
枯落物N元素含量为2.76%,P元素含量为0.22%,K元素含量为0.69%,木质素含量为6.83%,纤维素含量为 19.73%(表1)。
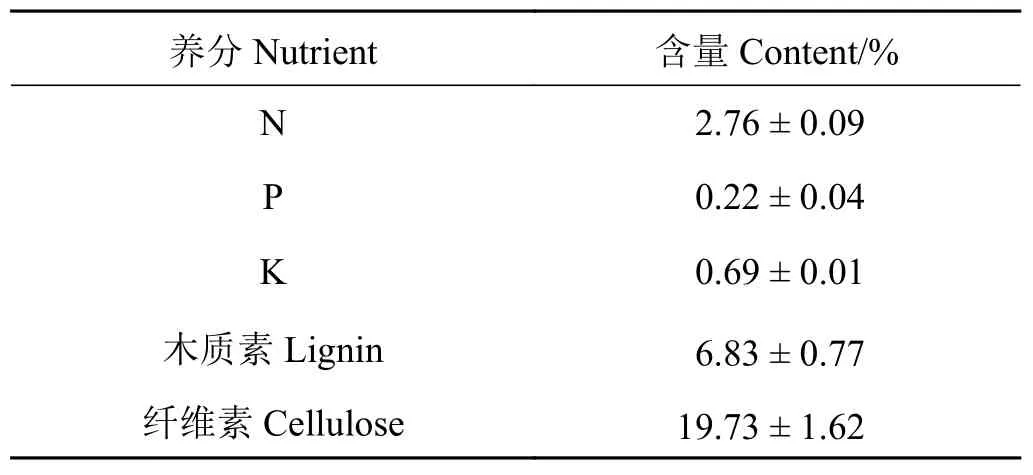
表1 柠条枯落物养分初始含量值Table 1 Litter nutrient concentration in the Caragana korshinskii shrub microhabitats
2.2 4 mm网孔内枯落物养分元素变化动态
分解至210 d时,N和纤维素残留率随灌丛大小变化呈现差异(表2)。N在4类灌丛微生境中的释放状态表现为Ⅳ类(-80.04%)释放速度最快,Ⅲ类(-24.46%)释放速度最慢,Ⅱ类(-47.29%)和Ⅰ类(-63.63%)释放速度居中。纤维素在4类灌丛微生境中的富集状态变化与N元素的释放规律变化相似,表现为Ⅳ类(142.81%)富集速度最快,Ⅲ类(38.97%)富集速度最慢,Ⅱ类(68.43%)和Ⅰ类(97.80%)富集速度居中。但是,P、K和木质素含量变化随灌丛大小变化并未呈现显著差异性(P>0.05)。
2.3 2 mm网孔内枯落物养分元素变化动态
K和纤维素残留率随灌丛大小变化呈现差异(表2),随着灌丛的增大,K呈释放状态,表现为Ⅰ(-36.32%) < Ⅱ(-17.23%) < Ⅲ(-13.93%) < Ⅳ(-3.04%)。纤维素在4类灌丛微生境中呈富集状态,表现为Ⅰ类(160.44%)富集速度最快,Ⅱ(109.40%)、Ⅳ类(107.81%)富集速度居中,而Ⅲ类(70.36%)富集速度最慢。
比较4和2 mm网孔间柠条灌丛微生境中枯落物养分元素残留率变化特征,发现在分解至210 d时,N、P、K、木质素和纤维素残留率在两种网孔间均呈现出相似的变化规律,即N、K和木质素在该阶段表现为释放状态,而P和纤维素则表现为富集状态。

表2 3种网孔内不同灌丛大小微生境中柠条枯落物养分元素残留率Table 2 Residual rate of litter elements in three mesh sizes among different stages of shrubs%
2.4 0.01mm网孔内枯落物养分元素变化动态
K和P残留率随灌丛大小变化呈现出不同差异(表2)。K在4类灌丛微生境中的释放状态表现为Ⅲ类(-27.73%)释放速度最快,Ⅰ类(-2.04%)释放速度最慢,而Ⅱ类(-19.94%)和Ⅳ类(-2.33%)释放速度居中。P在4类灌丛微生境中的富集状态表现为Ⅰ类(121.03%)富集速度显著(P< 0.05)快于Ⅱ类(29.01%)和Ⅳ类(43.11%),但均与Ⅲ类(74.79%)无显著差异 (P> 0.05)。
比较4、2和0.01 mm网孔间柠条灌丛微生境中枯落物养分元素残留率变化特征发现,在分解至210 d时,N、K在3种网孔内均呈现释放状态,而P和纤维素在3种网孔内均呈现富集状态。但是,在0.01 mm网孔内,木质素含量与其他4种元素变化差异较大,表现为大型灌丛微生境中木质素呈现出富集状态,而极小型、小型和中型灌丛微生境中木质素呈释放状态。
2.5 不同网孔中柠条枯落物养分元素变化动态
在4类大小柠条灌丛微生境中,3种网孔内枯落物N、K和木质素变化均表现为释放状态,而P和纤维素则均表现出富集状态(表2)。仅在Ⅰ类灌丛微生境中,N、P、K、木质素和纤维素残留率在3种网孔间均存在明显差异(P< 0.05)。其中,N、P和K在3种网孔间的残留率均表现为0.01 mm(-32.99%、121.03%和-2.04%) > 4 mm(-63.63%、14.31%和 -6.95%) > 2 mm(-75.47%、9.79%和-36.32%),木质素在3种网孔间的残留率表现为0.01 mm(-10.56%) > 2 mm(-46.18%) > 4 mm(-49.59%),纤维素在3种网孔间的残留率表现为2 mm(160.44%) > 4 mm(97.80%) > 0.01 mm(21.56%)。但是Ⅱ类、Ⅲ类和Ⅳ类灌丛微生境中枯落物养分元素残留率在3种网孔间均无显著差异(P> 0.05)。
2.6 枯落物元素残留率与其初始质量的相关性分析
在2 mm网孔内,N初始含量与木质素残留率间存在极显著正相关关系(P< 0.01)。P初始含量与P的残留率间存在显著负相关关系(P< 0.05)。K初始含量与K的残留率间存在显著负相关关系(P< 0.05)。
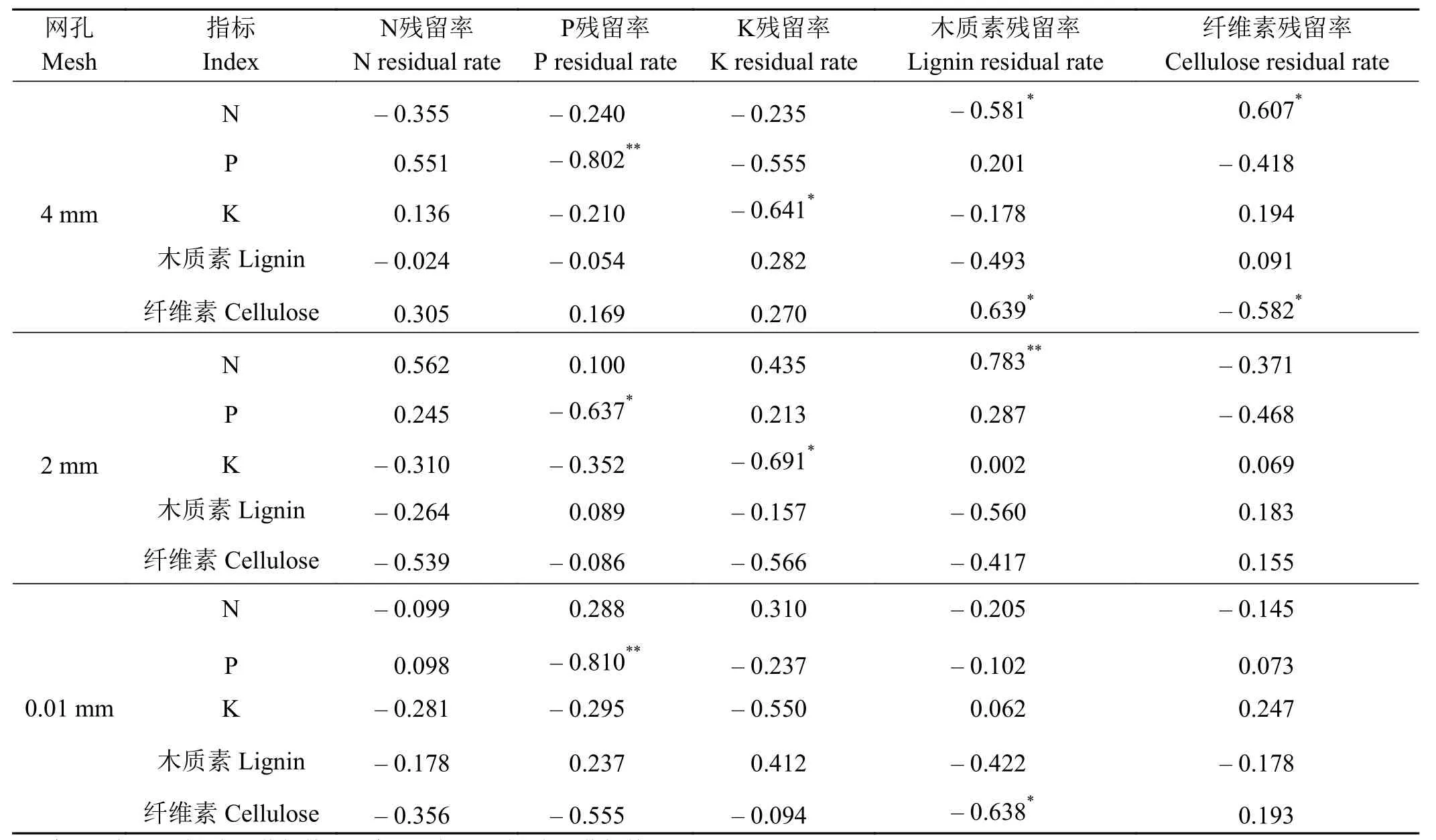
表3 枯落物元素残留率与其初始质量间的相关系数Table 3 Correlation coefficients between initial content of element residual rate and litter elements
在0.01 mm网孔内,P初始含量与P残留率间存在极显著负相关关系(P< 0.01)。纤维素初始含量与纤维素残留率间存在显著负相关关系(P< 0.05)。总体上看,灌丛微生境中枯落物N残留率与枯落物初始养分含量无相关关系(P> 0.05)。
不同大小柠条灌丛枯落物分解过程中元素残留率受枯落物初始养分含量影响程度不同(表3)。4 mm网孔内,N初始含量与木质素残留率间存在显著负相关关系(P< 0.05),而与纤维素残留率存在显著正相关关系(P< 0.05)。P初始含量与P残留率间存在极显著负相关关系(P< 0.01)。K初始含量与K残留率间亦存在显著负相关关系(P< 0.05)。纤维素初始含量与木质素残留率间存在显著正相关关系(P< 0.05),而与纤维素残留率间存在显著负相关关系 (P< 0.05)。
3 讨论
磷和氮是植物生长过程中最常见的限制性元素,对植物的各种功能影响深刻[20]。本研究中4类灌丛微生境中柠条枯落物分解至210 d时,在3种网孔内N元素均表现为释放状态,这与宁南山区草原柠条枯落物的研究结果一致[21]。并且,在不同大小柠条灌丛微生境中,N元素的释放均表现为较小型灌丛略高于较大型灌丛,这与黄土高原不同林龄柠条人工林的N含量变化相悖[11]。说明灌丛微生境的复杂程度会影响枯落物分解过程中养分的变化,同时受地理环境中温度与水分的影响而存在差异性。在4 mm网孔内,随着灌丛大小发生变化,N残存率间变化差异显著,而在2 mm网孔和0.01 mm网孔内,N元素残存率随灌丛大小变化差异不显著,主要是因为分解初期,参与分解的微生物会将分解释放的氮固定在其体内,也可能是枯落物在其分解过程中,微生物会从环境中吸收一些无机氮,进而调节C:N[22]。另外,分解初期的物理粉碎和淋溶过程也会使矿化出的N随着降雨而流失[23]。
在分解过程中,4类灌丛微生境中P呈富集状态,一般在枯落物分解初级阶段需要P元素富集,说明新鲜枯落物中的P养分的含量不足以维持分解者的生长,而枯落物分解速率与P元素及相关联的枯落物质量指标显著相关[24-26],表明枯落物的分解可能受P元素含量的限制。一方面,由于外源添加供给生态系统的P增加了枯落物中P的含量;另一方面,植物和微生物吸收的P在死亡后变成土壤有机磷,这部分P能够被重新矿化和吸收[27-28]。P元素受灌丛大小影响,在0.01 mm网孔内变化差异显著,而在4和2 mm网孔内随灌丛大小变化差异不显著,这与陈隆升等[12]的研究结果不一致。而马露莎[21]认为在柠条枯落物分解过程中,受土壤动物活动等影响,大、小孔径分解袋中P元素含量先减少再增加,最后大幅度减少,说明枯落物分解与底物组成有关,同时枯落物分解是一个长期的过程,本研究分析的枯落物分解至210 d时P元素的残存率变化规律存在一定的限制性。
在分解至210 d时,3种网孔内的枯落物K含量均呈现释放状态,是因为K元素主要以离子状态发生移动,因此表现为直接释放状态,这与相关研究[29-30]结果一致。在4 mm网孔内灌丛大小对枯落物K的影响不显著,而2和0.01 mm网孔内枯落物K元素变化受灌丛大小影响显著,同时在2和0.01 mm网孔内,随灌丛大小变化K的残存率变化规律不一致,这说明网孔大小影响枯落物分解的环境,进而影响枯落物K的变化。而且本研究结果与陈隆升等[12]对油茶枯落物的研究结果不一致,并未出现枯落物K含量成林大于幼林的现象。说明枯落物分解过程中K元素的变化既受灌丛大小的影响又受枯落物的底物质量影响。
木质素是枯落物的主要组成成分,其含量的多寡影响着枯落物的分解过程[31]。本研究发现,经过210 d的分解,4类灌丛微生境中枯落物在3种网袋内木质素多呈释放状态,这与对冰草(Agropyron cristatum)枯落物木质素含量的研究结果一致[32]。主要是因为研究期间为12月到次年7月,随着冻融期的结束,温度回升,土壤生物活动频繁,而土壤生物的活性增强有利于枯落物木质素结构的进一步物理破坏,因此使得木质素含量降低,这与前人的研究[33-35]结果一致。但是,在0.01 mm网孔中,大型灌丛柠条枯落物木质素含量呈现富集状态,说明在枯落物分解过程中,其组分与微生物作用形成了类似于木质素的酸不溶组分,进而导致枯落物木质素的增加[36],另一方面,枯落物中易分解组分(可溶性碳、氮、磷等)在季节性降雨期随雨水淋溶损失,导致易分解组分在凋落物中所占比例迅速降低,促使木质素等难分解组分所占比例有所提高。本研究结果显示,柠条灌丛大小并未对木质素含量产生显著影响,可能是因为只有特定的微生物才能将木质素分解,并且这种特定的微生物丰富度与灌丛微生境无关[32]。
纤维素是地球上含量最为丰富的天然高分子聚合物,也是植物残体中最丰富的部分,在植物细胞壁中普遍存在。已有研究表明,纤维素的分解受环境中纤维素酶的控制[37],同时,还会因枯落物的种类和植被类型的不同而受到影响[38]。本研究结果显示,3种网孔分解袋内的枯落物在分解至210 d时,纤维素呈富集状态,这与马志良等[35]的研究结果相悖,而与对黄土丘陵区草地生态系统的典型植被枯落物研究[32]结果一致,主要是因为随着分解的进行,纤维素受木质素的包裹和共同阻碍[39],在细胞壁中木质素与半纤维素通过多种键进行共价联结,包裹并保护植物细胞壁中的纤维素,形成纤维素衍生物,因此纤维素含量会增加并开始富集[32]。在4和2 mm网孔内,纤维素变化规律受灌丛大小影响显著,而0.01 mm网孔内枯落物纤维素变化不显著,说明随着柠条灌丛的发育,大、中型灌丛更有利于土壤有机质和营养物质的富集而形成丰富的土壤“生育岛”或“资源岛”,进而影响土壤动物类群活动[40],加剧分解者对枯落物的分解作用。
本研究结果显示,枯落物分解过程中养分元素随柠条灌丛大小变化规律并不明显,这与相关研究[11-12]结果存在一定的差异。造成枯落物养分元素发生富集或者释放的原因相当复杂,而分解本身就是一个复杂、长期的生物化学过程,分解中产生的中间产物,有时被分解者所利用,有的重新合成新的化合物,有的通过淋溶作用直接进入周围环境[41]。因此,在一个复杂的基质内,不能将每种化学成分分开处理,总有一些物质分解的快,有一部分分解的慢,分解过程中,有些物质可能发生着相互变化[42]。
4 结论
1)在分解至210 d时,N、K在3种网孔内均呈现释放状态,而P和纤维素在3种网孔内均呈现富集状态。但在0.01 mm网孔内,大型灌丛微生境中木质素呈现出富集状态,而在极小型、小型和中型灌丛微生境均呈释放状态。
2)当允许土壤动物和微生物参与时,大型灌丛微生境中枯落物N元素释放速度最快,纤维素富集速度最快。当允许中小型土壤动物与微生物参与时,极小型灌丛微生境中枯落物K元素释放最快,而纤维素富集最快。当仅允许微生物参与时,中型灌丛微生境中K元素释放速度最快,而极小型灌丛微生境中P元素富集速度最快。
3)在宁夏荒漠草原地区,不同大小灌丛微生境中柠条枯落物N、K、P元素和纤维素变化规律均相似,受土壤动物类群分布的影响较小。但是,柠条枯落物木质素释放则受到土壤动物类群分布和柠条灌丛大小的双重影响。
