宁夏荒漠草原柠条锦鸡儿枯落物分解特征及其影响因素
2019-07-08刘佳楠常海涛张安宁罗雅曦刘任涛
刘佳楠,常海涛,赵 娟,张安宁,陈 蔚,张 静,罗雅曦,刘任涛,*
1 宁夏大学西北退化生态系统恢复与重建教育部重点实验室,银川 750021 2 宁夏大学农学院,银川 750021
枯落物分解是陆地生态系统中物质循环和能量流动的主要途径,通过枯落物分解释放到土壤中的营养元素可以增加土壤肥力,维持地球生物化学循环[1-2],是深刻理解陆地生态系统结构和功能、养分循环规律的重要途径[3]。在干旱荒漠草原区,柠条锦鸡儿(C.kornshinskii)是防风固沙造林的重要灌丛树种之一[4],这些灌丛能够形成“沃岛”[5],成为干旱生态系统中生物地球化学过程最活跃的地方[6]。其中,柠条锦鸡儿枯落物累积与分解是灌丛微生境中释放矿质养分供植物生长的重要土壤生态过程[7],对于灌丛“沃岛”形成和维持干旱区脆弱荒漠草原生态系统元素平衡具有重要意义[8]。并且,采用不同网孔的分解袋来完全或部分限制土壤动物进入,是目前研究不同类型无脊椎动物分解功能普遍采用的方法。因此,基于不同网孔分解袋法,研究荒漠草原灌丛枯落物分解特征及其土壤生物分解作用,对于揭示荒漠草原生态系统生物地球化学循环过程及机制、草地管理及恢复利用均具有重要意义。
已有研究结果表明,随着灌丛林龄增加和林地生长发育,“沃岛”对于灌丛间土壤-植被系统产生重要的辐射作用[9],结果使得单一的灌丛生态系统逐渐演变成为较为复杂的灌草复合生态系统[9-10]。但是,关于不同发育阶段灌丛微生境中枯落物分解特征及对灌“沃岛”形成的作用机理,尚不清楚。根据前期大量文献查阅表明,目前关于不同林龄植物枯落物分解特征的研究,主要集中在森林生态系统[11-13]。侯玲玲等[14]对不同林龄的天然次生白桦林植物枯落物分解特征进行研究,发现枯落物分解与林龄大小存在一定的关系,并且枯落物的分解均表现出明显的阶段性。王欣[15]和吕瑞恒[16]等分别对华北落叶松人工林和油松人工林叶凋落物的分解特征进行了分析,发现叶凋落物的分解率随林龄的增加而增大。综合分析表明,不同林龄反映了不同阶段森林生态系统演变,林地发育过程变化对植物枯落物分解率、分解特性及分解系数均产生深刻影响。但是,在宁夏荒漠草原,关于不同发育阶段柠条锦鸡儿枯落物分解率变化特征及其影响因素研究,报道较少。
鉴于此,本研究以宁夏荒漠草原区柠条锦鸡儿枯落物为研究对象,采用不同网孔分解袋法研究不同发育阶段灌丛微生境中枯落物分解率及其分解过程,旨在揭示荒漠草原区柠条锦鸡儿枯落物分解过程对草地养分循环的影响,为荒漠草原生态系统生物地球化学循环过程解析、人工林建设与沙漠化防治提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于宁夏回族自治区盐池县,该区地处内蒙古自治区、甘肃省和陕西省的交界处,位于毛乌素沙地南缘,总面积71.3万km2,地理坐标为106°59′—107°07′E,37°56′—38°02′N,平均海拔1400 m。该区域属典型温带大陆性季风气候区,晴天多,降雨少,光能丰富,日照充足,年平均气温7.89℃。年降水量250—350 mm,其中7—9月占全年降水量的62%。年均无霜期为165 d,年平均风速为4.89 m/s。研究样地为该县域东北方向10km处(107°27′E,37°49′N,海拔1416 m)。土壤表面为风沙土覆盖,且粗砂粒含量超过90%,土壤肥力低下。主要植物有柠条锦鸡儿(C.kornshinskii)、油蒿(Artemisiaordosica)、花棒(Hedysarumscoparium)、赖草(Leymussecalinus)、胡枝子(Lespedezabicolor)、猪毛蒿(Artemisiascoparia)、牛枝子(Lespedezapotaninii)和沙蓬(Agriophyllumsquarrosum)等。
1.2 研究方法
1.2.1试验设计与分解袋布设
于2016年10月,在研究样地内选取26株柠条锦鸡儿灌丛进行标记,测量指标包括高度(cm)、冠层最大直径(cm)和最小直径(cm)。然后,根据灌丛大小(高度 × 冠幅),选取生长健壮、旺盛的灌丛样株进行标记,分成极小型(71 cm × 115 cm)、小型(122 cm × 519 cm)、中型(155 cm × 145 cm)、大型(223 × 341 cm)4类,分别记为Ⅰ、Ⅱ、Ⅲ、Ⅳ,代表了不同发育阶段的柠条锦鸡儿灌丛微生境演变过程。每类大小灌丛包括3个重复(即3株灌丛生长健旺、高度相近标记为样株),共有12个(4类 × 3重复)标记灌丛样株。
在标记灌丛样株下的东、南、西、北4个方向布设样方,收集地表枯落物,然后根据地表柠条锦鸡儿枯落物分布特征,计算单位面积柠条锦鸡儿枯落物干物质量,为5.8 g/cm2。然后,在相同生境中收集新鲜柠条锦鸡儿叶片(带叶柄),带回实验室自然风干后,依据分解袋规格(15 × 15 cm2)称取6 g分别装入4 mm(允许所有土壤动物和微生物进入)、2 mm(允许中小型土壤动物和微生物进入)、0.01 mm(允许微生物进入)网孔的尼龙网袋中。在分解袋上标好编码,并做记录。枯落物分解袋由两层尼龙网缝合而成,贴地面层均采用0.01 mm网孔的尼龙网,避免枯落物损失和地下土壤动物的干扰。
在极小型、小型、中型、大型4类灌丛微生境中布设上述3种规格的枯落物分解袋。从灌丛下正东方向开始布设,每个灌丛下依次均匀布设分解袋12个(3种规格 × 4次取样),分解袋距灌丛根部15 cm,且每两个分解袋间间隔10 cm。然后,用铁丝弯勾固定分解袋,使分解袋与地表土壤紧密接触,埋藏深度为0—2 cm。在2016年12月初完成分解袋布设,并且分别于0、120、150、210 d取回枯落物样品进行分析处理。共布设分解袋144个,即:3种规格 × 3个重复 × 4种类型灌丛 × 4次取样=144个。
1.2.2枯落物取样与分析
每次取样时,按照同样的顺序从每个灌丛下取回样品3袋,即每次取回36袋样品(3种规格 × 3个重复 × 4种类型灌丛)。将取回的样品装入信封,保存在恒温箱内,带回实验室。首先清除枯落物表面附着的泥沙和其他杂质,然后将取回的枯落物从分解网袋中转移到干净的信封中,置于烘箱,于70 ℃烘干48 h,称重,并记录枯落物样品的剩余干重。
1.2.3土壤样品取样与分析
枯落物分解袋取出后,在每个分解袋贴地表处,用铝盒取土壤剖面10 cm深度处的土壤,以测定土壤含水量;然后,用小铲将分解袋贴地表处其余土壤(0—10 cm)取回放在自封袋中带回实验室,自然风干,过2 mm土壤筛,去除树根等杂质,测定土壤pH和电导率。
土壤含水量(%)采用烘干称重法进行测定。土壤pH(2.5∶1水土比悬液)和电导率(5∶1水土比浸提液;μs/cm)分别采用酸度计法和电极法进行测定。土壤温度(℃)采用地温计连续3天进行测定,取平均值。
1.2.4数据处理与统计分析
枯落物分解率计算公式:
D=(M0-Mt)/M0× 100%
为了比较不同生境枯落物分解率的差异,本文参考了Olson衰减指数模型计算分解常数k,并根据此模型推算出了枯落物分解50%和95%所需要的时间T0.5和T0.95[17]。Olson衰减指数模型计算公式为:
Mt=M0e-kt
式中,Mt为枯落物在t时刻的质量(g),M0为初始质量(g),t为分解时间,k为枯落物分解常数。
采用单因素方差分析(One-way ANOVA),分析比较不同数据组间的差异,采用皮尔森相关系数(Pearson)分析数据组间的关联程度。所有统计方法采用SPSS 15.0软件完成,作图用Excel 2016完成。
2 结果与分析
2.1 土壤环境特征
从图1可以看出,不同大小灌丛下土壤含水量随时间变化均呈现出显著改变(P<0.05)。Ⅰ类、Ⅱ类和Ⅳ类灌丛下土壤含水量均表现为120 d显著高于150 d(P<0.05),而Ⅲ类灌丛下土壤含水量则表现为0 d和120 d显著高于150 d和210 d(P<0.05)。4类大小灌丛下土壤温度均表现为随时间变化呈显著增加趋势(P<0.05),即0 d<120 d<150 d<210 d。
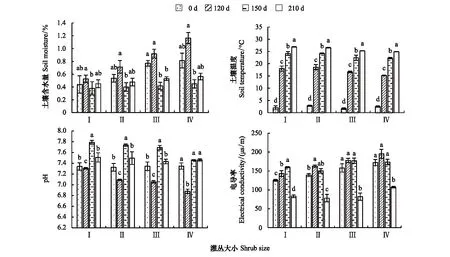
图1 柠条锦鸡儿灌丛微生境土壤性质变化Fig.1 Changes of soil properties in C.kornshinskii shrub microhabitats Ⅰ、Ⅱ、Ⅲ、Ⅳ 分别表示极小型、小型、中型、大型柠条锦鸡儿灌丛;不同小写字母表示同一类型灌丛下不同分解阶段间土壤性质差异性显著(P <0.05)
Ⅰ类、Ⅱ类和Ⅲ类灌丛下土壤pH值随时间变化均呈现相似变化特征,表现为150 d时最高,120 d时最低,0 d和210 d时居中。这与Ⅳ类灌丛下土壤pH值随时间变化分布特征差异较大。Ⅳ类灌丛下土壤pH值随时间变化表现为0、150、210 d时显著高于120 d时(P<0.05)。灌丛大小对土壤电导率随时间的分布特征亦影响较大。Ⅰ类和Ⅱ类灌丛下土壤电导率表现为从0 d到150 d时逐渐增加,而随后210 d时显著下降(P<0.05);Ⅲ类和Ⅳ类灌丛下土壤电导率均表现为0 d—150 d时显著高于210 d时(P<0.05),而0 d、120 d和150 d时3个时间点间无显著差异性。
2.2 基于灌丛大小的柠条锦鸡儿枯落物分解率
由图2可知,在4 mm网孔中,分解至120 d时,4类大小灌丛微生境中枯落物分解率变化表现为Ⅱ(41.07%)>Ⅰ(40.95%)>Ⅳ(39.65%)>Ⅲ(38.35%);分解至150 d和210 d时,4类大小灌丛微生境中枯落物分解率变化均表现为Ⅱ(41.73%,46.81%)>Ⅲ(38.92%,39.17%)>Ⅰ(35.20%,38.51%)>Ⅳ(33.76%,35.96%)。但是,在120 d、150 d和210 d时,4类大小灌丛微生境间枯落物分解率均无显著差异性(P>0.05)。
在2 mm网孔中,分解至120 d时,4类大小灌丛微生境中枯落物分解率变化表现为Ⅳ(36.65%)>Ⅱ(34.75%)>Ⅲ(33.41%)>Ⅰ(32.14%);分解至150 d,4类大小灌丛微生境中枯落物分解率变化表现为Ⅳ(38.33%)>Ⅰ(37.64%)>Ⅱ(34.34%)>Ⅲ(32.72%);分解至210 d,4类大小灌丛微生境中枯落物分解率变化表现为Ⅳ(48.05%)>Ⅰ(39.91%)>Ⅲ(39.01%)>Ⅱ(26.59%)。但是,在120 d、150 d和210 d时,4类大小灌丛微生境间枯落物分解率均无显著差异性(P>0.05)。
在0.01 mm网孔中,分解至120 d时,4类大小灌丛微生境中枯落物分解率变化表现为Ⅱ(46.81%)>Ⅲ(39.17%)>Ⅰ(38.51%)>Ⅳ(35.96%);分解至150 d,4类大小灌丛微生境中枯落物分解率变化表现为Ⅱ(38.25%)>Ⅲ(31.20%)>Ⅳ(26.88%)>Ⅰ(26.68%);分解至210 d,4类大小灌丛微生枯落物分解率变化表现为Ⅲ(39.90%)>Ⅰ(34.41%)>Ⅱ(33.80%)>Ⅳ(30.80%)。但是,在120 d、150 d和210 d时,4类大小灌丛微生境间枯落物分解率均无显著差异性(P>0.05)。

图2 3种网孔内不同大小柠条锦鸡儿枯落物分解率随时间变化Fig.2 Changes of litter decomposition rate with time for three-mesh litter bags between different shrub sizes 不同小写字母、带下划线的小写字母、带方框的小写字母、带方框和下划线的小写字母分别表示极小型、小型、中型、大型柠条锦鸡儿灌丛枯落物分解随时间的显著变化
2.3 基于分解袋网孔的柠条锦鸡儿枯落物分解率
由图3可知,在Ⅰ类灌丛微生境中,分解至120 d和150 d时,枯落物分解率在3种网孔间差异性显著(P<0.05);但在210 d时,枯落物分解率在3种网孔间无显著差异性(P>0.05)。表现为在120 d时,枯落物分解率4 mm(40.95%)和0.01 mm(38.51%)显著高于2 mm(32.14%);在150 d时,2 mm(37.64%)网孔内枯落物分解率最高,4 mm(35.20%)网孔内枯落物分解率次之,0.01 mm(26.68%)网孔内枯落物分解率最低。
在Ⅱ类灌丛微生境中,分解至120 d时,枯落物分解率在3种网孔间差异性显著(P<0.05),但在150 d和210 d时,灌丛微生境中枯落物分解率在3种网孔间差异性不显著(P>0.05)。具体表现为120 d时,4 mm(41.07%)网孔内枯落物分解率最高,0.01 mm(29.34%)网孔内枯落物分解率最低,2 mm(34.75%)网孔内枯落物分解率居中。
在Ⅲ类灌丛微生境中,在120 d、150 d和210 d时,枯落物分解率在3种网孔间均无显著差异性(P>0.05),这与其他3类灌丛微生境枯落物分解率变化差异较大。
在Ⅳ类灌丛微生境中,分解至120 d和210 d时,枯落物分解率在3种网孔间差异显著(P<0.05),但在150 d时,枯落物分解率在3种网孔间差异不显著(P>0.05)。具体表现为,120 d时4 mm(39.65%)网孔内枯落物分解率显著高于2 mm(36.65%)和0.01 mm(27.98%)网孔内枯落物分解率;210 d时,2 mm(48.05%)网孔内枯落物分解率最高,0.01 mm(30.80%)网孔内枯落物分解率最低,4 mm(35.96%)网孔内枯落物分解率居中。

图3 4类灌丛微生境中不同网孔分解袋枯落物分解率随时间的变化Fig.3 Changes of litter decomposition rate with time between three-mesh litter bags in four shrub microhabitats types 不同小写字母、带下划线的小写字母、带方框的小写字母分别表示表示4 mm、2 mm和0.01mm网孔中枯落物分解率不同分解阶段间的显著差异性;*表示相同时间点同种柠条锦鸡儿灌丛微生境中枯落物分解率在不同网孔间的显著差异性
2.4 枯落物残留率的Olson指数衰减回归分析
由表1可以看出,Olson指数能够较好的模拟宁夏荒漠草原柠条锦鸡儿枯落物分解动态(P<0.05)。在Ⅰ类和Ⅲ类灌丛微生境中,3种网孔内枯落物分解50%和95%所有用的时间一致;Ⅱ类灌丛微生境中,分解50%和95%所需的时间均为2 mm>0.01 mm>4 mm;Ⅳ类灌丛微生境中,枯落物分解50%和95%所需的时间均表现为0.01 mm>4 mm>2 mm。
不同大小柠条锦鸡儿灌丛微生境枯落物在同一网孔内分解50%和95%均呈现相出差异性。在4 mm网孔中,Ⅱ类灌丛微生境枯落物分解50%和95%所用时间最短,分别为0.63 a和2.74 a,其余3类分解时间相同;在2 mm网孔内,Ⅱ类灌丛微生境枯落物分解50%和95%所需时间最长,Ⅰ类和Ⅲ类中所需时间居中,Ⅳ类所需时间最短;在0.01 mm网孔内,Ⅳ类灌从微生境枯落物分解50%和95%所需时间均长于其他3类灌丛。

表1 柠条锦鸡儿枯落物残留率的Olson衰减指数模型Table 1 Decay exponential model of Olson in C.kornshinskii shrub microhabitats
Ⅰ、Ⅱ、Ⅲ、Ⅳ 分别表示极小型、小型、中型、大型柠条锦鸡儿灌丛
2.5 枯落物分解与环境因素的关系
由表2可知,不同发育阶段柠条锦鸡儿枯落物分解率受土壤环境因素的影响程度不同。土壤温度对柠条锦鸡儿枯落物分解快慢的影响达到显著正相关水平(P<0.05),同时对Ⅰ类、Ⅱ类、Ⅲ类灌丛微生境中枯落物分解产生显著影响(P<0.05),说明枯落物分解过程中,土壤温度是影响其分解快慢的重要指标。而土壤含水量和电导率对枯落物分解呈负相关关系,但未达到显著水平(P>0.05)。
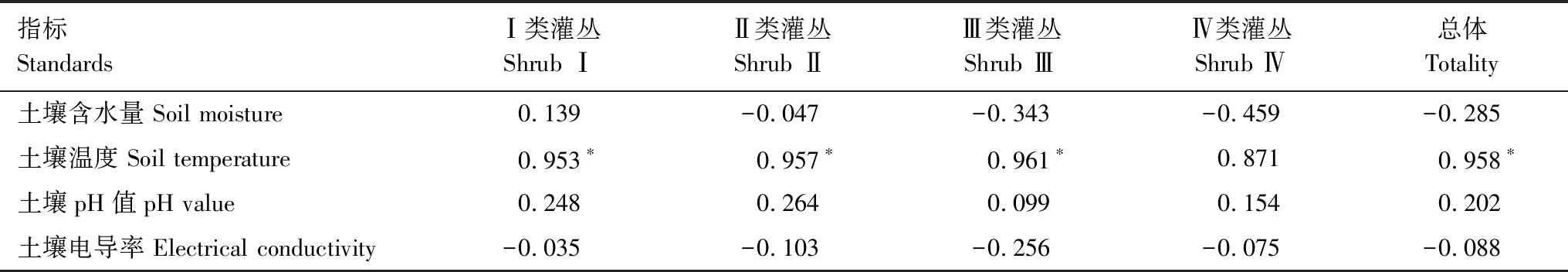
表2 枯落物分解率与土壤性质间的相关系数Table 2 Correlation coefficient between litter decomposition rate and soil properties
*表示在0.05水平(双侧)上显著相关.
3 讨论
3.1 不同大小柠条锦鸡儿灌丛对土壤性质的影响
一些研究表明,灌丛土壤含水量会随着植被林龄增加而降低[18-19]。而在荒漠草原区,由于土壤干旱,草本植物的竞争力远不如灌木,导致灌丛内草本植物较少,耗水量小;并且柠条锦鸡儿灌丛根系深广,可以吸收和消耗周围较大面积和深层土壤中的水分[20];加上微环境的影响,使土壤蒸发减少,导致柠条锦鸡儿灌丛土壤水分含量随着冠幅增大而增大,这与关林靖等[21]的研究结果一致。本研究中,大型、中型灌丛土壤含水量显著高于小型、极小型灌丛土壤含水量,这表明在荒漠草原区柠条锦鸡儿灌丛的发育增强了土壤的保水能力。另有研究表明,灌丛覆盖在影响土壤含水能力的同时会对土壤温度产生一定的影响[22]。研究中同一类型柠条锦鸡儿灌丛下土壤温度随时间延长显著升高,主要原因是该地区属于典型的温带大陆性季风气候,夏季高温,冬季寒冷,导致灌丛下土壤温度表现为0 d<120 d<150 d<210 d。
随着柠条锦鸡儿灌丛增大,土壤pH呈现一定的下降趋势(图1),这与靳虎甲等[23]在腾格里沙漠南缘古浪县东北部的鸣沙山和刘任涛等[24]在宁夏盐池县境内东北部区域研究结果一致。随着分解时间延长,同一灌丛土壤pH值在不同时间差异性显著。特别是在150 d时,土壤pH值显著高于其他时间土壤pH值,主要是因为柠条锦鸡儿为深根系灌木,其灌丛冠幅大、树干光滑、叶面积指数小,因此其冠层持水力较小[25-26]。降水在汇集入渗土壤之前,会在植物冠层有较长的接触滞留时间,而且树干组织具有较强的分泌能力[27],导致土壤pH值偏低。据气象数据显示,盐池县在12月—4月有不同程度的降水和降雪,而5月降水量偏低,因此在分解150 d左右,土壤pH值明显偏高。
随分解时间延长,同一发育阶段的柠条锦鸡儿灌丛土壤电导率呈增加趋势,这与Su和Zhao等[9]的研究结果吻合,而灌丛大小间变化规律与曹成有等[28]研究结果相悖。在柠条锦鸡儿灌丛发育过程中,根系有机酸的分泌以及和微生物的相互作用均可能导致土壤pH下降[9,29]。而土壤电导率的升高,可能与枯落物中可溶性盐的积累和沉积有关[29]。
3.2 不同大小柠条锦鸡儿灌丛微生境对枯落物分解的影响
枯落物分解率与枯落物质量、环境条件和分解者群落3个因素有关[30]。研究发现柠条灌丛大小对其微生境中枯落物分解率影响不显著(见图2),这与Su和Zhao等[9]的研究结果存在差异,并且逯军锋等[31]对不同林龄油松人工林凋落物进行研究,发现不同林龄枯落物分解率表现为成熟林>近熟林>中林龄>幼林龄,这与本文研究结果亦有区别,主要是因为本研究对象柠条灌丛间林龄差别小,时间序列短而导致不同大小灌丛间分解率变化差异小。不同大小柠条灌丛枯落物分解在各分解阶段表现出相似的变化规律,在分解过程中表现为快速分解(0—120 d)和缓慢分解(120—210 d)两个阶段[15]。在分解0—120 d时,灌丛微生境枯落物分解率上升较快,主要是因为在枯落物分解过程中,前期主要分解一些较易分解的物质,分解迅速。而在120 d后,枯落物分解率波动变化较小,因为在分解后期,主要分解一些难以分解的物质(木质素、纤维素等),因此分解率较慢。这与梁宏温[32]和邱尔发等[33]的研究结果一致。分解至120—150 d时,灌丛微生境中枯落物分解率在3种网孔内均开始下降,因为此时正值春季,温度回暖,土壤动物开始活动[34],并且受到昼夜温差和急速升温的影响,土壤动物回迁[35]以及刚从冬眠中苏醒的土壤动物急需摄入大量食物来保证生命活动,而土壤微生物是线虫、原生动物、变形虫和节肢动物等土壤动物的主要食物来源之一,是碎屑食物链的基本组成部分,一些土壤动物的捕食行为可能对土壤微生物产生一定的影响[36],进而导致微生物的量较少。但是在分解150—210 d时,枯落物分解率有呈现出平缓的上升趋势,这与林窗效应有关[37]。
网孔大小限制土壤动物的类群活动,进而对枯落物分解产生影响(见图3)。在分解至120 d时,大型和极小型灌丛微生境中4 mm网孔内枯落物分解率最高,而小型和中型灌丛微生境中则是0.01 mm网孔内就枯落物分解率最高,调查发现,枯落物中螨类和跳虫数量与该阶段枯落物分解率呈现正相关,这与余琴等[38]的研究结果吻合。在分解150 d和210 d时,极小型和大型灌丛微生境中枯落物分解率在4 mm网孔内最高,而极小型和大型灌丛微生境枯落物分解率则表现为2 mm网孔内最高。主要是因为在分解120 d后,土壤动物可对枯落物进行粉碎,增大枯落物的表面积,为微生物分解提供有利条件;同时,土壤动物的排泄物养分含量丰富,可以为微生物提供蛋白质和生长物质,刺激微生物生长,使枯落物更容易分解[39]。
经过Olson衰减指数模型分析发现(表4),在极小型和中型灌丛微生境中,柠条灌丛枯落物分解50%和95%均需0.95 a和4.1 a,而在小型灌丛和大型灌丛微生境中,枯落物分解50%需要0.63—1.90 a,分解95%需要2.74—8.21 a。综合来说,柠条锦鸡儿枯落物分解50%和95%所需的时间与蒙古栎阔叶混交林分解50%和95%所需时间相似[40],比不同林龄橡胶凋落物叶分解50%和95%所需时间长[41],却比华北落叶松人工林叶凋落物分解50%和95%所需时间短[15]。说明枯落物分解枯落物分解不仅受枯落物质量影响,亦与研究区内环境条件有关。
3.3 环境因子对枯落物分解的影响
温度是调控生化过程与物质能量周转的关键因子,同时也是影响枯落物分解的主要因素。研究发现,土壤温度对枯落物分解率变化影响显著(表2),短期内,温度升高可通过增加土壤微生物活性来促进枯落物分解,从而加快了草地生态系统中的物质循环;在长时间尺度方面,枯落物分解率随温度升高呈指数增加[42]。土壤含水量对枯落物分解存在抑制作用,但并未达到显著性水平。在干旱或半干旱生态系统中,水分因素可能成为限制枯落物分解的主要因子[43],即使在降水量高的地区,水分的季节性变化导致的阶段性干旱也会限制枯落物分解[44]。当降水量<100 mm时,土壤大部分时间处于极度干旱状态,枯落物分解受降水控制[45];当降水量>200 mm时,土壤含水量较低,但可维持土壤生物活性,脉冲式降水对枯落物分解的影响显著下降[46]。而土壤pH值与电导率对枯落物分解率的影响并不显著,主要是因为在局域尺度内,枯落物累积与分解除了受水、热因子限制,还受群落结构、土壤条件、植食动物等因素影响,表现较大变异性[47]。
4 结论
(1)随着分解时间延长,不同大小柠条锦鸡儿灌丛微生境土壤含水量和土壤温度均呈现出相似的分布特征,而土壤pH值和电导率随时间分布特征受到灌丛大小的显著影响。(2)在分解过程中,中型灌丛微生境中枯落物分解率受土壤动物类群分布影响较小,但其他类型灌丛微生境中枯落物分解率受土壤动物类群分布影响较大。(3)在宁夏荒漠草原,仅考虑柠条锦鸡儿灌丛大小时其微生境中枯落物分解率变化较小,而柠条锦鸡儿枯落物分解率受到灌丛大小和土壤动物类群相互作用的影响。
