杭州湾灰鳖洋海域秋季主要游泳动物生态位与种间联结
2019-07-08俞存根邓小艳颜文超
刘 惠,俞存根,郑 基,邓小艳,张 平,颜文超,谢 旭
浙江海洋大学水产学院,舟山 316022
生态位理论与种间联结性已成为现代生态学的重要内容[1],自Johnson最早使用生态位一词迄今已有百年历史[2]。美国学者Grinnell[3]首次将生态位概念运用于鸟类种群生态学研究,之后Elton[4]提出了功能生态位理论。20世纪50年代Hutchinson[5]在此基础上建立了多维超体积模式(n-dimensional hypervolume niche)并提出基础生态位和实际生态位的概念,随后生态位研究成为生态学研究的热点问题。经过不断完善和发展,该理论被广泛地接受,而我国是在20世纪80年代才对其全面展开研究。近30年该理论被越来越多的运用于群落结构及演替、生物多样性、种群进化与环境的关系等方面,并成为解释群落中物种关系的基础理论[6]。我国对生态位的研究主要集中在植物、昆虫、鱼类以及大型底栖动物[7]。另外,在生态学研究中,对种间关系的研究也同样重要。种间关系是指物种间在不同生境中相互吸引或排斥的性质[8]。我国对种间关系的研究主要集中在植物[9-13]、和鸟类[8,14-15]群落中,近几年才逐渐运用到水生生物方面[1,7,16-18]。将生态位和种间联结结合在一起,能够更加全面揭示群落中各物种对资源的利用程度。
目前,对游泳动物生态位和种间联结的研究中,针对不同季节的研究较多,如胡成业[7]、董静瑞[1]、张苗苗[18]等。然而,针对游泳动物在不同年份同一季节的相关变化的研究较为少见,仅看到对温州湾春季主要游泳动物的研究报道[16]。此类研究可以排除因季节变化造成生物资源的差异,因此在反映群落结构及其演替、生物多样性等方面具有一定的说服力。
杭州湾灰鳖洋海域位于舟山群岛和宁波之间,是舟山渔场重要的组成部分。该海域岛屿众多,水域地形复杂,受东海外海水和钱塘江径流以及水团的季节变化影响使得这一水域营养盐较为丰富,渔业资源多样性较高[19]。近年来对该海域水生生物的研究只出现在浮游动物[20],针对主要游泳动物的研究还未见报道,因此探究杭州湾口灰鳖洋海域主要游泳动物生态位与种间联结对了解当地的渔业资源现状具有十分重要的意义。本文通过对该海域的主要游泳动物的生态位和种间联结性去探索物种间的资源利用情况以及生态关系,以期为该海域的渔业资源利用和环境保护提供重要依据。
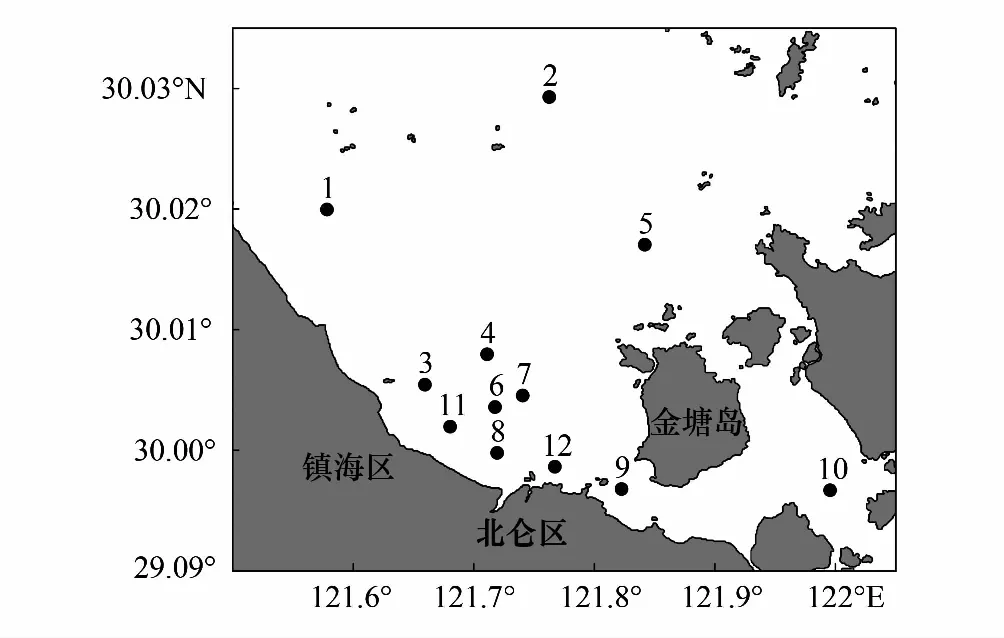
图1 杭州湾灰鳖洋海域渔业资源调查站位Fig.1 Survey stations of fisheries resource in Hui-bie-yang sea area of Hangzhou Bay
1 研究区域与方法
1.1 数据来源
数据取自2015年10月和2017年10月在杭州湾口灰鳖洋海域进行的底拖网渔业资源调查时所获得的游泳动物资源资料,调查共设12个站位(图1)。调查方法按照《海洋渔业资源调查规范》(SC/9403-2012)进行。网具规格两年分别为800目×160 mm,700目×70 mm,拖速约为3.5 kn,拖时约为0.5—1 h。对所获渔获物全部低温保存带回实验室分析鉴定。本文以胡成业[7]、邓小艳[17]、张苗苗等[18]相关研究为参考,对两年秋季计算得出的优势种、常见种进行筛选,将两个航次均出现的游泳动物组成的群体定义为主要游泳动物。
1.2 数据处理
由于本次调查航次在两个年份调查的网具规格和每站拖曳时间存在差异,为了方便比较分析灰鳖洋海域资源的分布状况,首先对所取渔获物数据进行标准化处理,然后选取两年秋季主要游泳动物的丰度和生物量数据进行分析。
1.2.1优势种测度
优势种采用Pinkas相对重要性指数[20]公式如下:
IRI=(N+W)×F
式中,N为某一物种个体数占渔获物总尾数的百分比;W为某一物种个体质量占渔获物总质量的百分比;F为某一物种出现的站位数占全部站位数的百分比。当IRI值大于1000时将该物种定义为优势种;当IRI值介于100—1000之间时,将该物种定义为常见种[21]。
1.2.2生态位测度
生态位宽度采用Shannon指数[22]:
生态位重叠值采用Pianka指数[23]:
式中,Bi指生态位宽度,其范围在[0,R]之间,值越大说明该物种生态位宽度越大;Pij与Pkj为种i和种k的尾数在j站位总尾数中的比值;R代表总站位数;Qik表示生态位重叠值,其范围在[0,1]之间。
1.2.3种间联结性测度
采用Schluter[24]提出的方差比率法(VR)来检验物间的总体关联性,以及用W值来检验关联显著性,公式如下:
VR=ST2/σT2
W=VR·n
式中,S为主要游泳动物全部种数,pi=ni/n,n为总站位数,ni为物种i出现的站位数;t为平均物种数,Tj为在站位j中出现的主要游泳动物的物种数。当VR为1时,表明物种间无关联;当VR大于1时,表明物种间存在正相关;当VR小于1时,表明物种间存在负相关。根据W值来判定VR值偏离1的显著性:若无关联,W值落在2分布的90%置信区间20.95(n) 根据连续校正公式[25]计算2值,公式如下: 式中,n为总站位数;a为种对间两物种同时出现的站位数;b和c为仅一种出现的站位数;d为种对间两物种均无出现的站位数。以ad-bc的正负性来判定物种间的正、负联结性。当2<3.841时,表明种对间独立;当3.841≤2<6.635时,表明种对间有显著联结性;当2≥6.635时,表明种对间有极显著的联结性。 联结系数(AC)[26]公式如下: 当ad≥bc时,AC=(ad-bc)/(a+b)(b+d) 当bc>ad且d≥a时,AC=(ad-bc)/(a+b)(a+c) 当bc>ad且d AC值的取值范围为[-1,1]。当AC值为0时,表明种对间完全独立;AC值越趋近于-1,表明负联结越强;AC值越趋近于1,表明种对间正联结越强。 共同出现百分率(PC)[26]公式如下: PC=a/(a+b+c) PC的取值范围为[0,1],其值越趋近于1,说明种对间正联结越紧密;为0时表明种对间无关联。 调查站位图采用Surfer 11软件绘制,数据处理分析应用Excel、Primer 5.0软件完成。 两个航次拖网调查共鉴定渔获物61种,2015年10月渔获物经分析共鉴定出生物种类43种,隶属于11目23科36属,2017年10月共鉴定出40种,隶属于10目23科35属。本文研究的主要游泳动物共21种,其中由相对重要性指数(IRI)公式计算得出,优势种有龙头鱼(Harpodonnehereus)和安氏白虾(Exopalaemonannandalei)2种,常见种有棘头梅童鱼(Collichthyslucidus)、葛氏长臂虾(Palaemongravieri)、脊尾白虾(Exopalaemoncarincauda)、三疣梭子蟹(Portunustrituberculatus)、日本蟳(Charybdisjaponica)5种(表1)。 表1 杭州湾灰鳖洋海域主要游泳动物生态位宽度及优势种Table 1 Niche breadth and dominance of major species of nekton in Hui-bie-yang sea area of Hangzhou Bay 杭州湾口灰鳖洋海域主要游泳动物的生态位宽度变化范围在0.01—3.92之间(表1),从中可以看出各游泳动物生态位宽度存在差异,基于生态位宽度可将其划分为3段,a段游泳动物的生态位宽度值较高,分布在(Bi≥1.0)范围内,为广生态位种;b段游泳动物生态位宽度值分布于(0.1≤Bi≤1.0)之间,为中生态位种;c段游泳动物生态位宽度值分布在(Bi≤0.1)范围内,为狭生态位种(图2)。 对该杭州湾灰鳖洋海域21种主要游泳动物生态位宽度进行聚类分析,可以划分成3组(图3)。第Ⅰ组由龙头鱼和安氏白虾组成。第Ⅱ组由棘头梅童鱼、六丝钝尾虾虎鱼、绒毛细足蟹、中国毛虾、葛氏长臂虾、三疣梭子蟹、日本蟳、口虾蛄、凤鲚、鲻和巨指长臂虾组成。第Ⅲ组由孔虾虎鱼、刀鲚、鮸鱼、红鳗虾虎鱼、中华栉孔虾虎鱼、日本鼓虾、拟穴青蟹组成。该聚类分析结果与生态位宽度分段结果呈现90.48%一致。生态位宽度值较高的物种有安氏白虾和龙头鱼,它们分布范围广且数量较大,而生态位宽度值较低的物种有鲻、红鳗虾虎鱼等,它们的分布范围窄且数量少。 图2 主要游泳动物生态位宽度分布Fig.2 Distribution of the niche breadth of major nekton species1.刀鲚 Coilia ectenes;2.凤鲚 Coilia mystus;3.红鳗虾虎鱼 Odontamblyopus rubicundus;4.棘头梅童鱼 Collichthys lucidus;5.孔虾虎鱼 Trpauchen vagina;6.六丝钝尾虾虎鱼 Amblychaeturichthys hexanema;7.龙头鱼 Harpodon nehereus;8.鮸鱼 Miichthys niiuy;9.中华栉孔虾虎鱼 Ctenotrypauchen chinensis;10.鲻 Mugil cephalus;11.安氏白虾 Exopalaemon annandalei;12.葛氏长臂虾 Palaemon gravieri;13.脊尾白虾 Exopalaemon carincauda;14.巨指长臂虾 Palaemon macrodactylus;15.日本鼓虾 Alpheus juponicus;16.中国毛虾 Acetes chinensis;17.拟穴青蟹 Scylla paramamosain;18.日本蟳 Charybdis japonica;19.绒毛细足蟹 Raphidopus ciliatus;20.三疣梭子蟹 Portunus trituberculatus;21.口虾蛄 Oratosquilla oratoria 杭州湾口灰鳖洋海域主要游泳动物生态位重叠值区间为[0—0.93](表2),重叠值高于0.85有7对依次是,1—5、20—21、18—21、11—12、2—9、7—20、2—4,表明这几种种类彼此之间利用资源的能力极具有相似性,其中重叠值为0的种对有3对,3—10、8—10、10—21,说明不同物种在同一生境中既相互重叠又相互独立,此外,优势种7(龙头鱼)分别与2、3、9、11、12、15、16、18、19、20、21号物种重叠值均超过0.6,重叠值偏高,说明优势物种与其他各主要游泳动物生态位重叠较为显著,种间竞争较强。 图3 主要游泳动物生态位宽度聚类分析Fig.3 Clustering analysis of niche breadth of major nekton species 根据方差比率可以求出VR=4.84,该值大于1,说明在独立零假设的情况下,主要游泳动物物种间关系总体上呈现正关联。根据21×24的物种-站位二元数据矩阵计算物种间的相关性(表3),通过统计量W来检验VR的显著程度,W值为58.03,不在置信区间[13.85,36.42]范围内,表明种间总体关联性显著。 基于卡方检验结果,在21种主要游泳动物组成的210个种对中,有9个种对呈极显著正关联,约占总种对数的4.29%;有105个种对呈现联结性显著,其中有21个种对呈显著负关联,约占总种对数的10%,还有38个种对呈显著正关联,约占种对数的18.1%;有37个种对呈极显著负关联,占总种对数的17.62%。在剩下的105个种对的联结性则表现为不显著,约占总种对数的50%。结果显示,安氏白虾、刀鲚、巨指长臂虾和三疣梭子蟹与其他主要游泳动物的联结性均呈现较高的水平(图4)。 表2 主要游泳动物生态位重叠值Table 2 Niche overlap of major species of netkon 表3 主要游泳动物种间的总体关联性检验Table 3 General interspecific association test of major netkon species 联结系数(AC)结果显示,种对间趋于独立的有42个,占总种对数的20%;种对间呈现为正联结的有95个,其中有34个种对间的联结性程度较高,占总种对数的16.19%,有61个种对间的联结程度一般,占总种对数的29.05%;种对间负联结程度一般的有14个,占总种对数的6.67%;有59个种对间的负联结程度较高,占总对数的28.1%(图5)。 共同出现百分率(PC)分析结果表明,正联结最强的种对有25个,占总种对数的11.9%;正联结性较强的种对有38个,占总种对数的18.1%;正联结性一般的有73个,占总种对数的34.76%;正联结性较弱的有49个,占总种对数的23.33%;正联结性最弱的有22个,占总种对数的10.48%;种对间无关联的有3个,占总种对数的1.43%。由此可见,种间联结性较强,总体趋于正相关(图6)。 生态位宽度是生物利用资源多样性的一个指标[27],游泳动物生态位宽度值的大小与其分布范围、分布格局以及种群数量有着密切的联系[7]。从生态学意义上说,生态位宽度又可以反映研究物种对外在环境的适应性以及利用资源的广泛性[28]。我们的研究发现主要游泳动物生态位宽度有较显著的分段情况,如图2所示,a段出现的物种为龙头鱼和安氏白虾,它们既是广生态位种也是优势种,在该海域的资源利用能力和环境适应性有较强的优势。b段中生态位种有葛氏长臂虾(0.86)、中国毛虾(0.51)、棘头梅童鱼(0.41)等10种主要游泳生物,该段物种所占比重最大达47.62%,且分布较为均匀。c段狭生态位种有中华栉孔虾虎鱼(0.07)、刀鲚(0.07)、巨指长臂虾(0.04)等9种主要游泳动物,它们所占的数量较少且分布的局限性较大,如巨指长臂虾在两年的调查中也仅在3、6、8、12号站位有分布。 图4 主要游泳动物种间2检验半矩阵图Fig.4 Semi-matrix diagram of the 2 test among the major nekton species 图5 主要游泳动物种间联结系数AC半矩阵图Fig.5 Semi-matrix diagram of inters pecific association (AC) among the major nekton species 图6 主要游泳动物种间共同出现百分率半矩阵图Fig.6 Semi-matrix diagram of the percentage co-occurrence (PC) among the major nekton species 本文调查的主要游泳动物的相对重要性指数(IRI)值与生态位宽度值虽然在广生态位种中无差异,但在中生态位种和狭生态位种中划分的序列并不完全一致,如中国毛虾生态位宽度值为(0.51)排第4位,而相对重要性指数值为(53.4)排第11位;绒毛细足蟹生态位宽度值为(0.18)排第9位,而相对重要性指数值为(11.4)排第16位,序列差异这可能是两者体现生态特征的角度不同所导致的[7]。生态位宽度计算侧重于物种与站位之间的联系,而相对重要性指数计算则是忽略了站位间的差异,利用该物种的尾数和生物量及出现频数来计算,因此生态位宽度与相对重要性指数序列会存在差异。 生态位重叠反映种对间利用资源能力所表现的相似性,它与物种分布站位点的重叠情况紧密相关,同时也反映出物种间的竞争关系,但是重叠值高并不意味着种间竞争一定激烈,重叠值是种间竞争的必要非充分条件[28]。在本文研究结果中,主要游泳动物生态位分布并不均匀,刀鲚和孔虾虎鱼重叠值最高(0.93),根据食性来看,刀鲚主要摄食浮游甲壳动物、鱼类和虾类[29],孔虾虎鱼以桡足类、枝角类、轮虫等浮游生物为食,在海洋生态系统中它们属于不同的营养级,此外,二者对栖息地的环境要求有所不同,尽管生态位重叠值较高,但并不意味着种间竞争一定激烈。 一般而言,种群的生态位宽度越宽,其他种群与其生态位重叠的可能性就越大,反之亦然。然而,本研究的结果发现,具有较大生态位宽度种对之间的生态位重叠值不一定很高。如中国毛虾生态位宽度(0.51)但与其他物种间的生态位重叠值较低,这种现象可能是由于物种本身生态学特性不同,对资源段位要求不完全一致,从而导致生态位重叠不高。 本研究应用种间联结性分析的方法对杭州湾南岸灰鳖洋海域底层游泳动物进行分析,并判断该区域群落所处的状态、演替的趋势和运动。这对研究群落的动态变化和组成具有一定的帮助[30]。经过方差比率与卡方检验对种间联结进行测定,我们得出物种间总体呈现正相关。这表明该水域的群落结构相对稳定,生态系统结构也较为成熟[16]。 卡方检验结果显示在210个种对间有105个种对表现为不显著。这可能是各个物种在该区域的相互依赖程度低所致。在姚琪[31]对杭州湾地区地质地貌的研究中,镇海金塘港区附近海域海底底质有厚大松散的堆积层,呈现二元结构沉积旋回的叠覆。这种海底地形可能会阻碍底层游泳生物之间的交流,促使它们各自占据有利生境,从而降低彼此的关联度。此外,也无法排除是否受到了人为因素的干扰,从而造成的群落演替程度上升,种对间关联度下降[32]。当然,2检验以二元数据矩阵作为判定标准,对种间相关的多度信息无法全面表达,因此只能作定性判断[10]。同时,卡方检验可以判断种对间联结性是否显著,并不能体现其大小,并且经2检验不显著的种对,也不意味着它们之间不存在联结性[11]。 PC和AC可以反映那些由2检验认为不显著的种对间联结性的大小[12]。因此,本文在2检验的基础上结合PC和AC对物种间联结性进行测定,使我们的研究结果更具说服力。AC分析显示,正联结种对所占比重最大,且相应的生态位重叠值也较高,说明群落间趋于稳定共存,群落越发成熟。但数据显示负联结性种对也占据一定比例,这可能与调查样方的面积有关[13]。当所研究群落中物种数量很大而调查样方面积较小时,结果只能反映出较少的物种数。因此,样方的科学选取,数量尺度的把握都是今后研究中应解决的问题[33]。根据PC值可知,在最强正联结种对中,日本蟳、三疣梭子蟹和龙头鱼与其他种对所占比重较大,这说明这些物种与其他游泳动物相比具备较强的竞争力和生存能力。但具备强正联结种对不一定生态位重叠值高,如日本蟳和三疣梭子蟹,这可能是与种对分布的频率、生境差异、食性以及对资源的利用等情况有关[17],具体原因还待进一步研究。 在杭州湾灰鳖洋海域秋季主要游泳动物中,龙头鱼和安氏白虾在该海域的资源利用能力和环境适应性有较强的优势,而中华栉孔虾虎鱼、刀鲚、巨指长臂虾等9种主要游泳生物,它们的所占的数量较少且分布的局限性较大。种间联结性分析结果表明该水域的群落结构相对稳定,生态系统结构也较为成熟。在2检验的基础上结合PC和AC对物种间联结性的测定结果显示正联结所占比重最大,且相应的生态位重叠值也较高,这表明虽然该水域受到诸多的人为因素干扰,但群落整体上仍趋于稳定共存,这也支持了种间联结性分析结果。在最强正联结种对中,日本蟳、三疣梭子蟹和龙头鱼与其他种对所占比重较大,说明了这些物种与其他游泳动物相比具备较强的竞争力和生存能力。不过,由于随时空尺度变化,物种的生境状况会发生改变,在群落中的关系及相对地位也会有所改变,这可能会影响到研究结果。在以后的研究中,应考虑将时空尺度的变化及东南沿海一些重要的自然生态现象(如赤潮、台风等)与相关生物资源数据进行耦合。 致谢:菅康康同学参与海上样品采集,韩磊、钱金生、戴冬旭、孙蓓蓓、辛艺、张文俊等同学参与实验室内种类鉴定,特此致谢。2 结果与分析
2.1 优势种

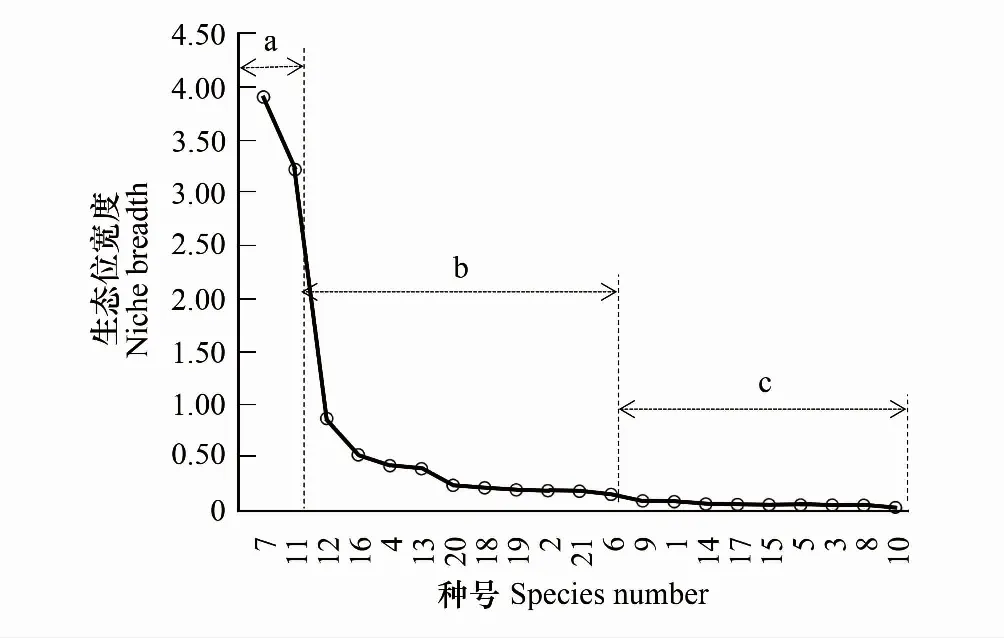
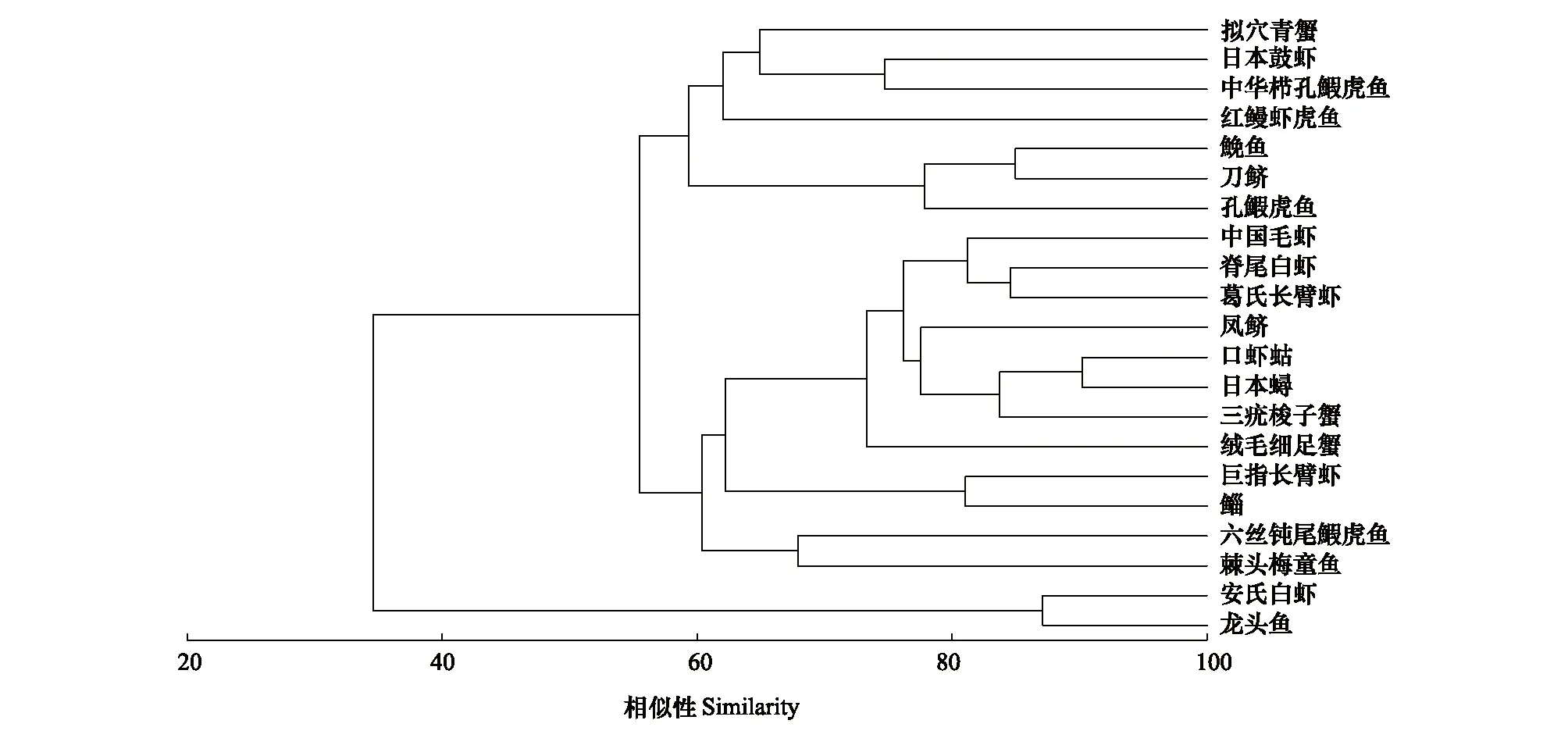
2.2 生态位宽度
2.3 生态位重叠
2.4 总体相关性
2.5 种间联结性


3 讨论
3.1 生态位宽度与生态位重叠特征

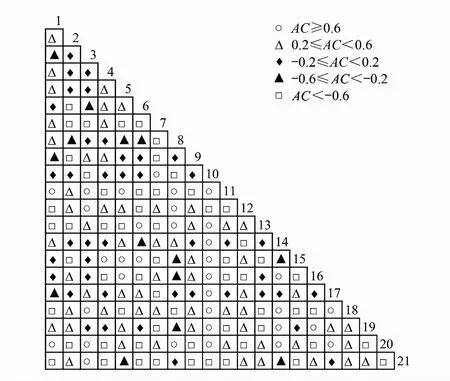

3.2 种间联结性分析
4 小结
