猪繁殖与呼吸综合征病毒Nsp2蛋白纳米抗体的筛选及其鉴定
2019-07-08孙明霞温永俊刘永刚蔡雪辉
宋 欢,孙明霞,尹 坤,温永俊,刘永刚,蔡雪辉*
(1.中国农业科学院哈尔滨兽医研究所,黑龙江哈尔滨150069;2.中国农业科学院特产研究所,吉林长春130112;3.内蒙古农业大学,内蒙古呼和浩特010018)
猪繁殖与呼吸综合征(Porcine reproductive and respiratory syndrome,PRRS)是 由 PRRS 病 毒(PRRS virus,PRRSV)引起的病毒性传染病,该病在临床上主要引起母猪流产以及各年龄段猪的呼吸系统疾病,给养猪业造成不可挽回的经济损失[1]。由于PRRSV 具有高度变异性和抗体依赖的增强作用,现有的疫苗并不能给猪群提供完全的免疫保护[2],因此有必要尝试研究新型抗病毒策略。PRRSV 的非结构蛋白 2 (Nonstructural protein 2,Nsp2)是 PRRSV 最大的非结构蛋白,它的顺式结构和反式结构均具有蛋白酶活性,对病毒的毒力至关重要,是研发抗PRRSV 药物的理想靶标蛋白[3]。
纳米抗体(Nanobody)具有分子量小、结构稳定、可溶性好、特异性强、生产成本低,以及可以识别特殊抗原表位等优点[4]。目前已有针对多种人类病毒纳米抗体的相关报道,研究结果显示其具有良好的临床应用前景,例如针对呼吸道合胞病毒的ALX-0171,是第一个处于临床阶段的抗病毒纳米抗体药物[5]。但在动物病毒疾病中纳米抗体鲜见报道。因此本研究纯化了PRRSV Nsp2 重组蛋白免疫骆驼,利用噬菌体展示技术筛选针对PRRSV Nsp2 的特异性纳米抗体,并对其进行鉴定,为后续PRRSV Nsp2 的研究提供物质基础,为研发新型抗PRRSV药物提供了实验依据。
1 材料与方法
1.1 质粒、菌株和实验动物 pET30a-Nsp2 质粒、鼠抗骆驼IgG 抗血清、PCV2 Cap 重组蛋白和PRRSV Nsp1α 重组蛋白由本实验室制备保存;BL21(DE3)感受态细胞购自Thermo 公司;pCANTAB 5E噬菌体、E.coliTG1 和辅助噬菌体M13KO7 均由西北农林科技大学周恩民教授实验室赠送;6~8 周龄BALB/c 小鼠购自北京实验动物中心;成年大棕双峰骆驼购自内蒙古农业大学。
1.2 主要试剂 PRRSV 抗体阳性猪血清由本实验室保存;鼠源anti-His 单克隆抗体(MAb)购自Life Technology 公司;山羊抗小鼠IgG-HRP 和山羊抗兔IgG-HRP 购自北京中杉金桥公司;Ni-NTA Agarose购自GE 公司;兔抗E-tag 多克隆抗体、Protein G 纯化介质购自南京金斯瑞公司;HRP 标记的鼠抗M13噬菌体 MAb 购自 Sino Biological 公司;RNeasyPlus Mini RNA kit 购自 QIAGEN 公司;Ficoll-PaquePLUS 淋巴细胞分离液购自GE 公司;Leucosep 淋巴细胞分离管购自Greiner bio-one 公司。
1.3 PRRSV Nsp2 重组蛋白在大肠杆菌中的表达及纯化 将pET30a-Nsp2 质粒转化BL21(DE3)感受态细胞,培养至OD600nm值0.6~0.8 时,利用IPTG(终浓度 1 mmol/L) 16 ℃ 180 r/min 诱导 16 h 后, 以 anti-His MAb 为一抗,羊抗小鼠IgG-HRP 为二抗,利用western blot 检测重组蛋白表达情况;同时,以PRRSV 抗体阳性猪血清为一抗,羊抗猪IgG-HRP 为二抗,利用western blot 检测重组蛋白的抗原性。鉴定正确后经过Ni-NTA 亲和层析纯化,参照试剂盒说明书测定纯化蛋白浓度。
1.4 双峰驼免疫及Nsp2 抗体效价测定 将5 mL Nsp2 重组蛋白(1 mg/mL)与等体积弗氏完全佐剂混合并充分乳化,颈部皮下免疫雄性大棕双峰驼,此后每隔两周用弗氏不完全佐剂与抗原充分乳化,使用同样方法加强免疫5 次,第3 次加强免疫后4 d采血分离血清,以Nsp2 重组蛋白为包被抗原,骆驼血清为一抗,鼠抗骆驼IgG 抗血清为中间抗体,羊抗鼠IgG-HRP 为二抗,参照文献[6]中ELISA 方法检测骆驼血清中Nsp2 抗体效价。免疫前血清作为阴性对照。若样品孔/ 阴性孔OD450nm值大于2.1判定为阳性。
1.5 VHH 噬菌体抗体文库的构建 最后一次双峰驼免疫后4 d,采集200 mL 抗凝血并用等体积RPMI1640 稀释,利用Ficoll-PaquePLUS 淋巴细胞分离液分离淋巴细胞,血细胞计数板计数后按1×107个细胞/ 支分装,使用RNeasyPlus Mini RNA kit提取RNA,反转录得到cDNA。根据参考文献[7]及双峰驼VHH 上下游基因序列,利用Primer Premier 5.0 软件设计引物(表1),由库美生物公司合成。

表1 VHH 片段扩增所用引物Table 1 The primers for amplification of VHH fragment
参照文献[6]套式PCR 方法进行文库鉴定筛选。第一轮PCR 以cDNA 为模板,利用引物CALL001/CALL002 进行。第二轮PCR 扩增以第一轮PCR 胶回收产物为模板,利用引物VHH-FOR/VHH-REV 扩增VHH 基因片段,胶回收目的片段后测定浓度,用于下一步实验。将VHH 基因片段通过PstⅠ和NotⅠ酶切位点插入pCANTAB 5E 噬菌体展示载体,将连接产物电转化TG1 感受态细胞,均匀涂布于LB/AMP-GLU 平板,收集菌苔,-80 ℃保存。取100 μL电转产物,用LB/AMP-GLU 培养基10 倍倍比稀释,各浓度分别取 100 μL 涂布于 LB/AMP-GLU 平板,37 ℃培养8 h,进行平板计数。随机挑选50 个单克隆进行菌液PCR 鉴定,鉴定为阳性的克隆进行测序分析,利用MegAlign 软件比对测序结果,鉴定文库多样性。
1.6 Nsp2 特异性重组噬菌体的淘选和富集 取100 μL 噬菌体展示载体文库培养至对数期,加入M13KO7 辅助噬菌体感染,过夜培养后离心收集上清,利用PEG/NaCl 方法纯化重组噬菌体颗粒,将重组噬菌体10 倍倍比稀释后侵染对数期的TG1 细胞,涂布于LB/AMP-GLU 平板计数菌落,计算重组噬菌体滴度。以Nsp2 重组蛋白包被酶标板,用2 %脱脂乳粉稀释重组噬菌体溶液至5×1011pfu/mL,加入酶标板,室温孵育2 h 后,弃去噬菌体样品,洗涤,每孔加入100 μL 新鲜配制的0.1 mol/L 三乙胺,室温静置10 min,吸出洗脱液迅速用等体积1 mol/L Tris-HCl (pH7.4)中和,测定洗脱液噬菌体滴度;富增洗脱的噬菌体用于下一轮筛选;重复上述步骤2次,完成第二轮和第三轮淘选。同时设PBS 包被酶标板作为无抗原对照。
将Nsp2 重组蛋白作为包被抗原,按照文献[8]中噬菌体ELISA 方法检测特异性重组噬菌体的富集情况。
1.7 重组纳米抗体的诱导表达及ELISA 检测
1.7.1 重组纳米抗体的诱导表达 取第三轮筛选后得到的噬菌体,10 倍倍比稀释后与对数期TG1 细胞混匀静置15 min,涂布于LB/AMP-GLU 平板,37 ℃培养8 h。随机挑选96 个单克隆分别用1 mL TB 培养基重悬后接种于24 孔板,37 ℃220 r/min 培养至对数期,每孔加入IPTG(终浓度1mmol/L) ,16 ℃180 r/min 诱导16 h,离心弃上清,-20 ℃冷冻30 min后室温放置15 min,每孔加入500 μL PBS 重悬菌体,离心收集上清即为可溶性重组纳米抗体粗提物,用于下一步实验。
1.7.2 可溶性重组纳米抗体的ELISA 检测 将Nsp2 重组蛋白包被酶标板,以纳米抗体粗提物为一抗,兔抗E-tag 多克隆抗体为中间抗体,羊抗兔IgG-HRP 为二抗,参照文献[6]中间接ELISA 方法检测重组纳米抗体,同时设PBS 作为阴性对照。根据OD450nm值判定,若样品孔/ 阴性孔该值大于2.1 判定为阳性。
1.7.3 Nsp2 特异性纳米抗体的测序分析 选择1.7.2 中ELISA 鉴定为阳性的克隆由库美公司测序,采用MegAlign 软件比对测序结果,根据纳米抗体高变区序列进行分析。
1.8 Nsp2 纳米抗体的特异性试验 根据测序结果,采用1.7.2 中ELISA 方法检测阳性纳米抗体的特异性和效价。取Nsp1α 重组蛋白(同样带有His 标签,表达纯化系统同Nsp2 重组蛋白)作为对照组,与Nsp2 重组蛋白同时包被酶标板,ELISA 方法检测Nsp2 纳米抗体特异性;将阳性纳米抗体粗提物倍比稀释,测定Nsp2 纳米抗体效价。
2 结 果
2.1 Nsp2 重组蛋白在大肠杆菌中的表达纯化与抗原性检测 将pET30a-Nsp2 表达载体转化BL21 感受态细胞,以anti-His MAb 为一抗,western blot 检测结果显示,诱导后菌体在70 ku 处存在目的蛋白表达,大小与预期相符,表明Nsp2 重组蛋白正确表达;以PRRSV 抗体阳性猪血清为一抗,western blot 检测结果显示,PRRSV 阳性猪血清可以特异性识别该条带,表明该重组蛋白具有抗原性(图1)。经检测,Ni-NTA 亲和层析柱纯化的Nsp2 重组蛋白浓度为1.7 mg/mL,蛋白产量约为10 mg/L。

图1 Nsp2 重组蛋白的western blot 鉴定Fig.1 Identification of recombinant Nsp2 protein of PRRSV by western blot
2.2 VHH 噬菌体抗体文库的构建 采用纯化的Nsp2 重组蛋白免疫双峰驼,采用间接ELISA 方法检测第3 次加强免疫后抗体效价,结果显示免疫骆驼血清中Nsp2 特异性抗体的效价可以达到1∶256 000。最后一次免疫后4 d 采集抗凝血,从200 mL 血液样品中一共分离到2×108个淋巴细胞。提取细胞总RNA 反转录得到cDNA,利用套式PCR 经两轮扩增,最终得到约450 bp 的目的条带。回收目的条带,将其克隆入噬菌体展示载体pCANTAB 5E,转化TG1 感受态细胞,最终得到库容为6.5×107cfu 的VHH 噬菌体展示文库。对随机挑选的50 株单克隆进行菌液PCR 鉴定,结果显示,96 %的克隆含有目的基因的插入(图略)。阳性克隆测序结果显示,噬菌体抗体文库多样性良好,可进行下一步淘选工作。
2.3 Nsp2 特异性纳米抗体的淘选和富集 将Nsp2重组蛋白作为包被抗原淘选特异性纳米抗体,在淘选过程中,通过检测每轮洗脱液中重组噬菌体的滴度来评估特异性VHH 重组噬菌体的富集效果。结果显示,在3 轮筛选过程中,噬菌体回收率逐轮升高,第三轮筛选后,P/N 值可以达到2 200 (表2),表明特异性重组噬菌体得到富集。取每轮筛选后扩增的噬菌体溶液进行噬菌体ELISA 检测,结果显示,OD450nm值呈逐轮增加的趋势(图2),表明经过3轮筛选后,Nsp2 特异性噬菌体得到了富集,与表2中数据结果一致。

表2 筛选过程中特异性噬菌体的富集情况Table 2 Enrichment of phage particles carring Nsp2-specific Nbs

图2 噬菌体ELISA 检测筛选过程中特异性噬菌体的富集Fig.2 Detection of the enrichment of phage particles carring Nsp2-specific Nbs by phage ELISA
2.4 Nsp2 特异性纳米抗体的诱导表达及测序鉴定结果 从第三轮筛选后的细菌平板上随机挑取96个单克隆,经扩增诱导后制备可溶性重组纳米抗体粗提物,采用ELISA 检测,初步鉴定结果显示,选取的96 个单克隆中有93 个阳性克隆(OD450nm值大于PBS 对照3 倍以上判定为阳性) (图3),表明筛选到的特异性纳米抗体与Nsp2 反应性良好。将ELISA检测为阳性的93 个克隆进行测序分析,氨基酸序列进化树分析结果显示阳性克隆中一共包含44 株不同的纳米抗体(图略)。抽取序列差异较大的8 株抗体序列与人的抗体重链可变区(VH)进行序列比对,结果显示,所有序列在FR2 区均具有纳米抗体典型的亲水氨基酸的替换(图4)。表明筛选到的纳米抗体具有良好的水溶性。

图3 间接ELISA 方法检测重组纳米抗体与Nsp2 的反应性Fig.3 Analysis of the binding ability of the recombinant nanobodies against Nsp2 by indirect ELISA
2.5 Nsp2 纳米抗体的特异性试验结果 利用间接ELISA 方法对44 株纳米抗体的特异性进行了分析,结果显示,44 株纳米抗体均可以特异性结合Nsp2重组蛋白,与PRRSV Nsp1α 蛋白无交叉反应,具有高度特异性。图5A 中显示44 株纳米抗体均可以特异性结合Nsp2 重组蛋白,与Nsp1α 重组蛋白无交叉反应;为了进一步分析44 株纳米抗体对Nsp2 的结合能力,通过ELISA 检测了纳米抗体效价,同时选择一株Nsp2 阴性纳米抗体(Nb57)作为阴性对照,结果显示相对于其它纳米抗体Nb11、Nb12 和Nb60效价更高(图5B)。表明44 株纳米抗体均具有良好的特异性,且其中Nb11、Nb12 和Nb60 具有更强的结合力。

图4 8 株纳米抗体的氨基酸比对Fig.4 Amino acid sequence alignment of the 8 Nsp2-specific nanobodies
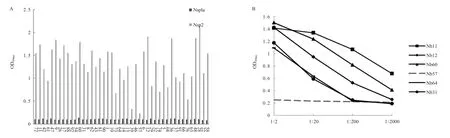
图5 ELISA 检测Nsp2 特异性纳米抗体的特异性(A)和结合力(B)Fig.5 Binding capacity (B) and specificity (A) of the nanobodies against Nsp2
3 讨 论
PRRS 给全球养猪业造成了严重的经济损失,如何对其进行防控一直是研究的热点。Nsp2 作为PRRSV 最大的非结构蛋白,在病毒的复制和毒力调控中发挥着举足轻重的作用。Nsp2 蛋白的N 端是一个木瓜样蛋白酶区(PL2),随后是一个包含丰富脯氨酸的B 细胞表位区和4 个预测的TM 单环的高变区,最后是一个保守的C 端[9];PL2 具有正向和反向切割蛋白酶活性,其正向切割活性是病毒复制过程所必需的;C 端可以引起膜结构重排,进一步形成双层囊泡,与多个病毒蛋白和宿主蛋白相互作用,参与病毒复制转录复合体的形成[10];Nsp2 蛋白由于含有大量的B 细胞表位,能够诱导高水平体液免疫反应,是一个潜在的良好诊断靶标[11];其还参与调节病毒RNA 诱导的干扰素的表达,具有免疫调节作用[12]。因此,本研究以Nsp2 为靶标蛋白,选取保守性较强的蛋白酶段和B 细胞表位段作为免疫原,淘选具有高亲和力的PRRSV Nsp2 纳米抗体,以期为PRRSV 的检测和防控提供新思路。
本研究选取纳米抗体而非单链抗体或Fab 段,是因为纳米抗体具有其它类型抗体所不具备的优势。本实验室先期纯化了骆驼IgG,SDS-PAGE 结果显示,双峰驼IgG 存在两种重链,一类分子量与传统IgG 重链分子量一致(55 ku 左右),另外一类分子量为43 ku 左右,与此前报道的HcAb 的重链大小一致[13]。本研究利用可溶性Nsp2 重组蛋白结合弗反佐剂免疫双峰驼,经过4 次免疫后,骆驼血液中Nsp2 特异性抗体的 ELISA 效价高达 1∶256 000,表明可溶性Nsp2 重组蛋白具有较好的免疫原性,机体对其产生了较强的体液免疫反应。
在构建VHH 噬菌体展示文库时,由于这类纳米抗体与人源VH 有很高的同源性,为避免VH 的干扰,本实验利用套式PCR 经两轮扩增后,将其克隆入pCANTAB 5E 载体,获得了库容为6.5×107cfu 的骆驼VHH 噬菌体展示文库,该库容量可以满足后续淘选实验,且序列分析结果表明文库具有较好的多样性。纳米抗体与传统的VH 最显著的区别在于其 FR2 区的 4 个氨基酸的替换(Val37Phe/Tyr、Gly44Glu/Gln、Leu45Arg 和 Trp47Gly/Phe/Leu,序号之前为VH 的保守氨基酸,之后为VHH 特有的亲水性氨基酸)[14],本实验淘选到了44 株纳米抗体,随机选取8 株,发现它们在FR2 区均存在典型的亲水氨基酸的替换,这些亲水氨基酸增加了VHH 的水溶性,对其溶解性和稳定性起到重要作用。此外,选取的8 株纳米抗体在CDR1 区和CDR3 区分别含有1 个半胱氨酸,研究显示,这对半胱氨酸可以形成二硫键,从而稳定抗原结合区的构象[15]。
在特异性试验中筛选到的Nsp2 特异性纳米抗体与同样带有His 标签的PRRSV Nsp1α 蛋白、PRV gE蛋白和PCV2 cap 蛋白均不能发生反应,表明筛选到的纳米抗体具有高度的特异性,可以满足其作为诊断和治疗用抗体的要求;同时检测了44 株特异性纳米抗体的效价,结果显示Nb11、Nb12 和Nb60 与Nsp2 重组蛋白具有更强的结合力。本实验采用噬菌体展示技术筛选到44 株PRRSV Nsp2 特异性纳米抗体,不仅可为Nsp2 蛋白的研究提供物质基础,还可以通过后续标记建立检测方法或进行抗病毒研究,应用于PRRS 的检测或防控。
