青海省森林细根生物量及其影响因子
2019-07-05唐立涛字洪标阿的鲁骥王长庭
唐立涛,字洪标,胡 雷,阿的鲁骥,王长庭
西南民族大学青藏高原研究院, 成都 610041
细根(直径≤2 mm)具有直径小、无木质部、生长和周转迅速的特点,是构成森林生态系统地下碳库的重要部分[1]。虽然森林细根生物量在大部分陆地生态系统地下总生物量中占比为3%—30%,但是它在维持自身功能的过程中最高能消耗净初级生产力的75%[2]。因此,探究细根动态过程对于了解细根在森林生态系统中碳分配和养分循环过程中的作用至关重要。但由于对根系的生态学作用认识不足以及根系研究的困难,人们研究重点往往是林木的地上部分,而对林木地下部分的研究相对较弱。
近年来,随着全球碳循环研究的开展以及对森林生态系统研究的深入,森林细根生物量变化规律受到了国内外学者的广泛关注。以往研究表明细根生物量不一定呈现水平分布格局[3],且不同海拔植被细根生物量及各形态指标都具有明显的垂直变化规律[4]。细根的分布与土壤容重、碳和氮含量有显著的相关性,土层深度也是影响细根生物量的重要因素[5]。鉴于细根对于森林生态系统的重要性,研究细根生物量对森林地下碳库的贡献不仅体现在局域水平,而且区域尺度上研究细根生物量对碳收支的贡献率也必不可少。然而,青海省森林作为青藏高原的重要组成部分,其对森林生态系统结构和功能的作用,及其与土壤物质循环(如碳氮循环)和维持机制仍不明确。
青海森林生态系统在固碳造氧、涵养水源、森林憩游、保育土壤以及生物多样性保护等方面贡献巨大,生态地位十分重要[6]。圆柏(Sabinachinensis)、云杉(Piceaasperata)、白桦(Betulaplatyphylla)等是其生态系统植被碳库的重要组成部分。当前针对青海云杉林研究所涉及的方面颇多,例如:青海云杉与其林下植物多样性的关系[7],云杉林更新的时空特征[8]。但有关青海省森林地下细根生物量及其影响因子等方面研究相对匮乏。因此本文选取云杉、白桦、圆柏、山杨(Populusdavidiana)和白杨(Populustomentosa)5种林分类型为研究对象,旨在分析和探讨青海森林细根生物量的分布格局及其与环境因子的关系,以期更深入地了解青海省森林生态系统碳汇以及地下碳循环过程,为该地区森林生态系统可持续利用与管理提供基础资料。
1 材料与方法
1.1 研究区域概况
青海省雄踞于青藏高原东北部(31°39′—39°19′ N,89°35′—103°4′E),是黄河、长江、澜沧江的发源地。青海省东西长约1200 km,南北宽约800 km,占地面积约72.1×104km2。其中林地面积为11.2×106hm2,占全省总面积的15.6%,以寒温带针叶林为主,且大多集中在海拔2000—4000 m区域[9]。地貌复杂,气候独特,属典型高原大陆性气候,年均气温-3.7—6.0℃,年日照2340—3550 h,年降水量16.7—776.1 mm(大部分400 mm以下),年蒸发量1118.4—3536.2 mm(大部分1500 mm以上)[10]。
1.2 研究方法
1.2.1样地设置与细根的采集处理
本研究依托中国科学院战略性先导科技专项(碳专项),按照《生态系统固碳现状、速率、机制和潜力》项目[11]制定的统一要求,并结合青海省森林资源连续清查成果,充分考虑全省各森林类型(优势种)分布面积、蓄积比重、起源等情况,在全省21个县(图1)布置主要森林类型标准样地80个,海拔为2175—3852 m,以垂直距离300 m分为6个梯度(表1)。每个样地随机设置3块50 m×20 m的乔木样方,各样方间距大于100 m,共计240个样方。每个样方按照平均木的标准选取优势种,利用内径5 cm的根钻,在优势树种树冠下随机钻10个土芯,按0—20 cm、20—40 cm分层取样。将样品用清水浸泡1 d,在流水中过100目土壤筛分离土壤和细根。根据细根的颜色、外形、弹性、皮层保留直径≤2 mm的木本植物细根。洗净后将细根称重,后分别装入纸袋,在85℃的条件下烘干并称重。
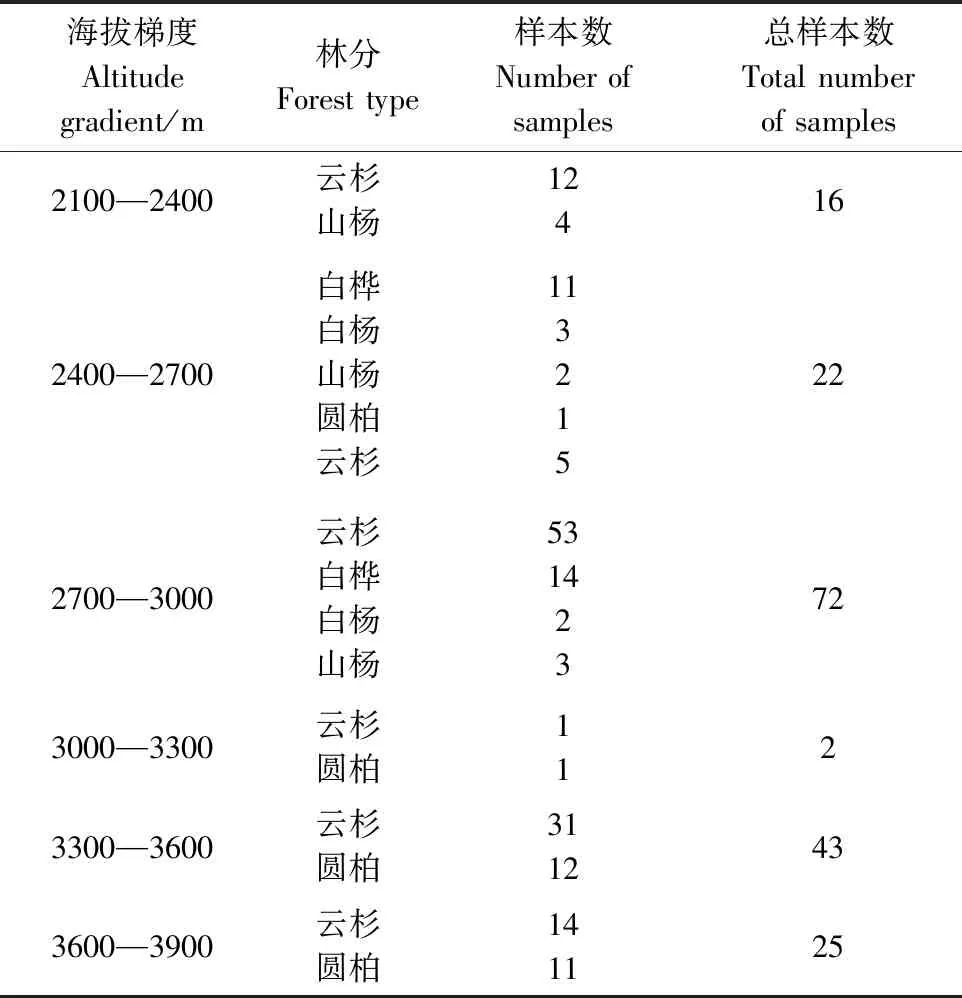
表1 不同海拔梯度样本分布
本文最终筛选出180个样本数据
1.2.2土壤理化性质测定
采集根系的同时,在每个样方随机选取样点并清理林地表层凋落物,采用土钻法(直径5 cm)采集0—20 cm和20—40 cm土层土壤各9个,每个土层的9个土样充分混合均匀,装入封口袋。将土样过1 mm筛清理土样中石块、残留根系及其他杂质后测定土壤容重和土壤养分TC、TN、TP含量。采用重铬酸钾容量法-外加热法测定土壤TC含量;凯氏定氮法测定土壤TN含量;钼锑抗比色法测定土壤TP含量。
1.3 数据处理
采用多因素方差分析(Multi-way ANOVA)检验土层和海拔、土层跟林分以及海拔和林分的交互作用对细根生物量的影响;单因素方差分析(One-way ANOVA)检验不同海拔、林分细根生物量的差异性,如果方差为齐性,采用LSD法进行多重比较,若方差为非齐性,则用Tamhane′s 2法进行多重比较;利用Pearson相关分析海拔、土层、土壤容重、土壤养分与细根生物量之间的相关性,显著性水平设置为α=0.05,以上分析均使用SPSS 10.0统计软件完成。利用AMOS 17.0构建细根生物量与环境因子的结构方程模型。运用Excel 2010软件完成数据的计算和图表绘制。
图1 青海省采样点位图Fig.1 Sampling point map of Qinghai Province
2 结果与分析
2.1 细根生物量的变化特征
2.1.1不同海拔梯度细根生物量的变化
土层、海拔、土层和海拔的交互作用均对青海省森林细根生物量有显著影响(P<0.05)(表2),细根生物量随海拔梯度总体上先降低再升高(图2)。细根生物量在海拔2100—2400 m处最大(10.27 t/hm2),且显著大于2400—3000 m和3300—3600 m处(P<0.05),在海拔2400—2700 m最小(7.65 t/hm2)。
同一土层细根生物量随海拔梯度的变化特征不规律,但差异较大(图3)。表层(0—20 cm)细根生物量在海拔2100—2400 m最高(8.01 t/hm2),3300—3600 m最低(5.42 t/hm2)。深层(20—40 cm)细根生物量在海拔3000—3300 m最大(3.04 t/hm2),2700—3000 m最低(1.99 t/hm2)。同一海拔不同土层,细根生物量随土层的加深明显下降,6个海拔梯度不同土层的细根生物量均差异显著(P<0.05)且表层细根生物量占比为68%—78%。
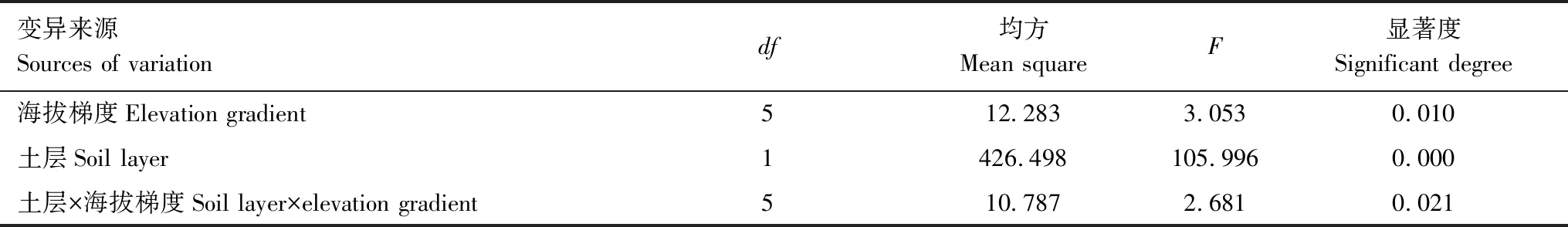
表2 海拔和土层对细根生物量影响方差分析
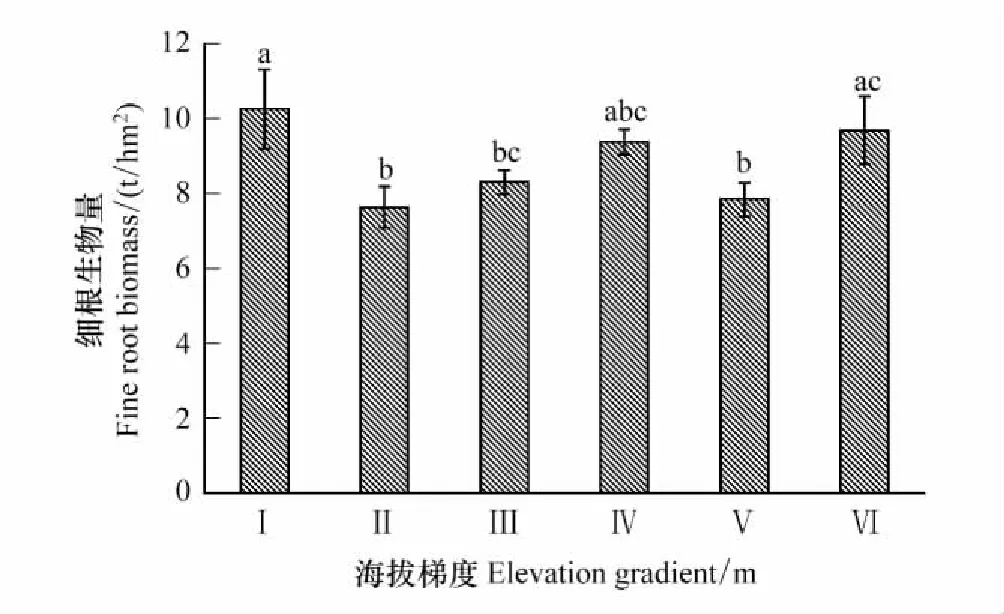
图2 不同海拔梯度细根总生物量的分布特征Fig.2 Distribution characteristics of the total biomass of fine roots at different altitudes横坐标中的Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ分别表示海拔梯度2100—2400 m、2400—2700 m、2700—3000 m、3000—3300 m、3300—3600 m、3600—3900 m,下同;不同小写字母表示不同海拔梯度细根生物量差异显著(P<0.05)
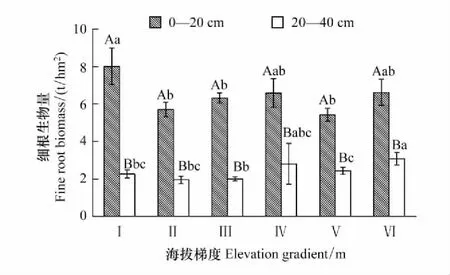
图3 海拔梯度对细根生物量的影响Fig.3 Fine root biomass at different altitudes不同小写字母表示同一土层不同海拔之间差异显著;不同大写字母表示同一海拔不同土层之间差异显著(P<0.05)
2.1.2不同林分细根生物量的变化
土层对不同林分细根生物量影响显著(P<0.05),而林分、林分与土层、海拔梯度和林分的交互作用对细根生物量影响并不显著(表3,表4)。5种林分细根总生物量大小为:白桦<白杨<云杉<圆柏<山杨(图4)。山杨林细根生物量最高(8.94 t/hm2),白桦林最低(7.75 t/hm2)。相同土层细根生物量在不同林分间差异不显著(图5),所有林分表层细根生物量大小为:白桦<圆柏<白杨<云杉<山杨,深层为:白杨<白桦<山杨<圆柏<云杉。
所有林分细根生物量的垂直分布特征大致相等,均随土层加深而减少。不同林分的表层细根生物量均显著大于深层(P<0.05),且表层细根生物量占总细根生物量的72%—80%。

图4 不同林分间细根总生物量的分布特征 Fig.4 The distribution characteristics of the total biomass of fine roots between different forest types不同小写字母表示不同林分细根生物量差异显著(P<0.05);BP:白桦Betula platyphylla;PT:白杨Populus tomentosa;PA:云杉Picea asperata;SC:圆柏Sebina chinensis;PD:山杨Populus davidiana
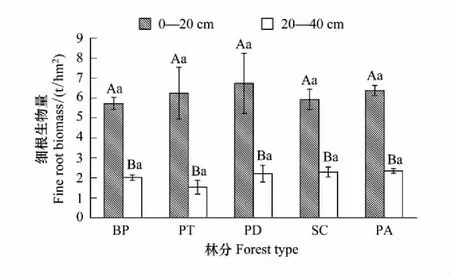
图5 林分对细根生物量的影响Fig.5 Effect of forest type on fine root biomass 不同小写字母表示同一土层不同林分之间差异显著;不同大写字母表示同一林分不同土层之间差异显著(P<0.05)

表3 林分和土层对细根生物量影响方差分析
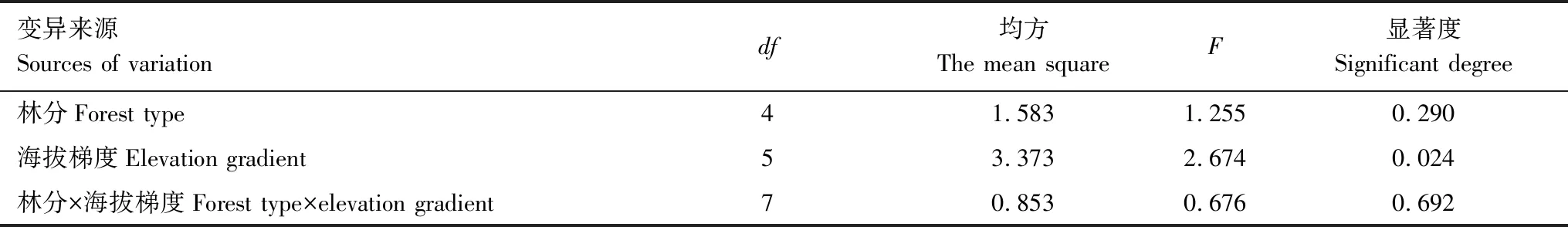
表4 林分和海拔梯度对细根生物量交互影响方差分析
2.2 土壤理化性质的变化特征
2.2.1不同海拔梯度土壤理化性质的变化
土壤容重大致随海拔增加而变大,最小值出现在2100—2400 m处,最大值出现在3300—3600 m处。土壤表层、深层土容重在低海拔处与高海拔处差异显著(P<0.05)。所有海拔梯度土壤容重均呈现出随土层加深而变大的垂直分布规律,且深层显著大于表层(P<0.05)(表5)。
各个海拔梯度土壤表层TC、TN、TP含量显著大于深层(P<0.05)。TC含量均随海拔梯度总体上先增加后减少,在3300—3600 m处达到峰值。TN含量随海拔梯度也表现出与TC类似的规律。TP含量随海拔梯度逐步减少,不同海拔上TP含量均存在差异,最小值出现在3600—3900 m处。
2.2.2不同林分土壤理化性质变化
不同林分土壤容重均存在差异,云杉、白桦林土壤容重较小,且与白杨、山杨林有较大差异(表6)。所有林分土壤容重均随土层的加深而变大。同一林分不同土层,深层(20—40 cm)土壤容重显著大于表层(0—20 cm)(P<0.05)。
所有林分TC、TN、TP含量均随土层加深而降低,且表层含量均显著大于深层(P<0.05)。云杉林土壤TC、TN、TP含量较高。表层土壤中,白杨林TC含量显著小于白桦、云杉林(P<0.05),云杉林TN含量显著大于白杨林(P<0.05),云杉、白桦林TP含量显著大于山杨、圆柏林(P<0.05)。深层土壤中,白杨、山杨林TC含量与白桦林差异显著(P<0.05),山杨林与圆柏、云杉林TN含量差异显著(P<0.05),云杉林TP含量显著大于圆柏林(P<0.05)。
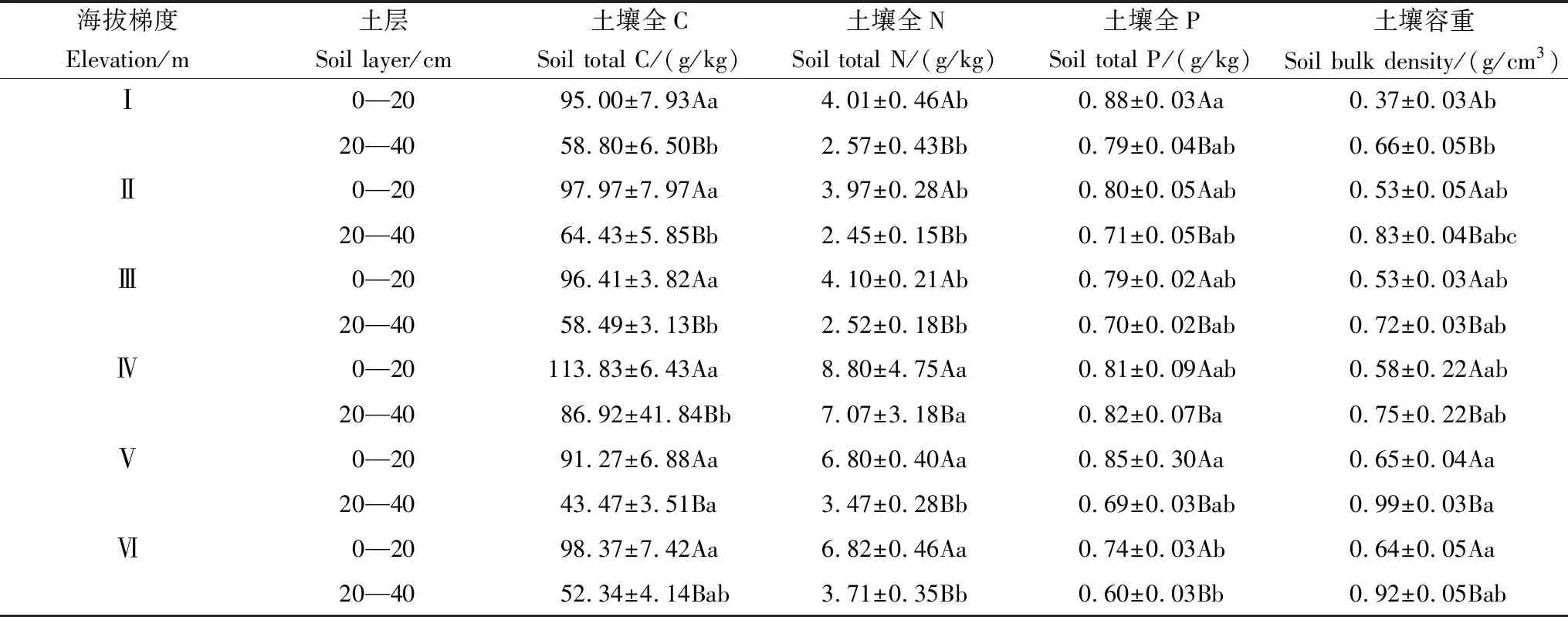
表5 不同海拔梯度内土壤特性分布特征
第一列中的Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ分别表示海拔梯度2100—2400 m、2400—2700 m、2700—3000 m、3000—3300 m、3300—3600 m、3600—3900 m,下同;不同小写字母表示不同海拔梯度相同土层细根生物量差异显著(P<0.05),不同的大写字母表示同一海拔梯度内不同土层间细根生物量差异显著(P<0.05)
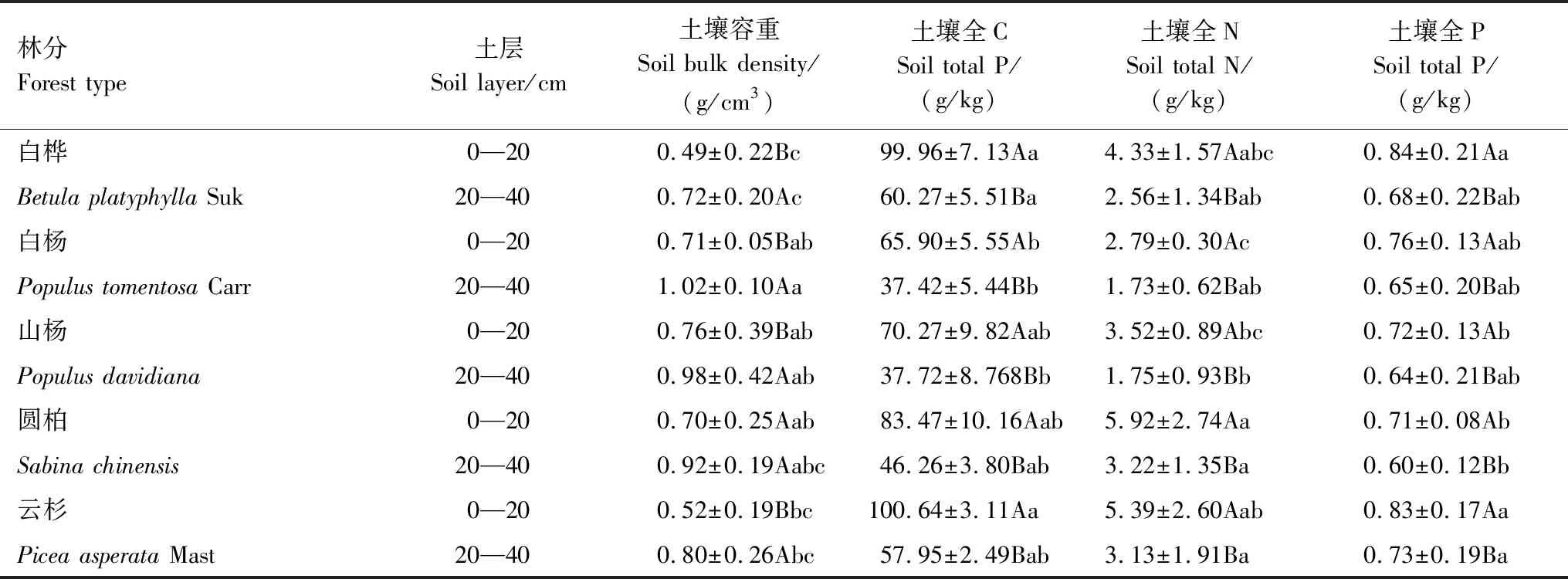
表6 不同林分土壤理化性质分布特征
不同小写字母表示不同林分间相同土层细根生物量差异显著,不同大写字母表示同一林分不同土层间细根生物量差异显著(P<0.05)
2.3 细根生物量与环境因子相关性分析
构建结构方程模型(Structural Equation Model, SEM)分析细根生物量与环境因子之间的相互影响关系。CHI/DF = 0.218,GFI = 0.999,RMSEA < 0.001,P= 0.929(图6)说明该模型能够较好的适配研究数据以及路径分析体系。建模时对土壤TC、TN、TP进行降维处理,提取第一维数据可解释率为63.62%。结果显示SEM能解释细根生物量的49%,土壤养分的48%,土壤容重的37%。土层、土壤容重直接影响细根生物量,并且极显著负相关(P<0.01);林分通过容重间接影响细根生物量;细根生物量显著影响土壤养分(P<0.01)。Pearson相关分析结果也表明:细根生物量与海拔、土壤TC、TN、容重、土层均极显著相关(P<0.01)。而土壤TC、TN、TP均与容重呈负相关(P<0.01)(表7)。
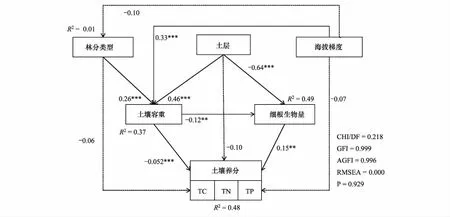
图6 细根生物量与环境因子结构方程模型Fig.6 The structural equation model of fine root biomass and environmental factor** P < 0.01,*** P < 0.001;实线表示影响显著,虚线表示影响不显著;CHI:卡方值,Chi-Square;DF:自由度,Degrees of freedom;GFI:适配度指数,Goodness-of-fit index;AGFI:调整后适配度指数,Adjusted goodness-of-fit index;RMSEA:近似误差均方根,Root mean square error of approximation
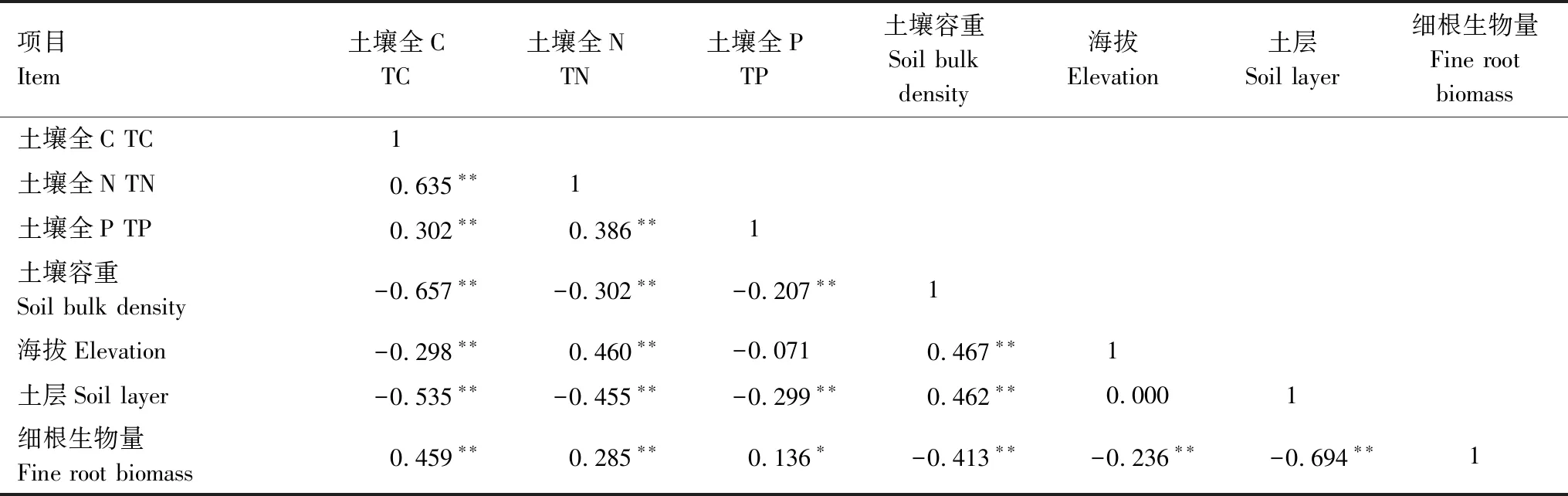
项目Item土壤全CTC土壤全NTN土壤全PTP土壤容重Soil bulk density海拔Elevation土层Soil layer细根生物量Fine root biomass土壤全C TC1土壤全N TN0.635∗∗1土壤全P TP0.302∗∗0.386∗∗1土壤容重Soil bulk density-0.657∗∗-0.302∗∗-0.207∗∗1海拔Elevation-0.298∗∗0.460∗∗-0.0710.467∗∗1土层Soil layer-0.535∗∗-0.455∗∗-0.299∗∗0.462∗∗0.0001细根生物量Fine root biomass0.459∗∗0.285∗∗0.136∗-0.413∗∗-0.236∗∗-0.694∗∗1
*P<0.05,**P<0.01
3 讨论
3.1 细根生物量的变化
研究表明,细根生物量随海拔梯度先降低后升高,在2100—2400 m处最大,因为低海拔处生境适宜,种群通过增加细根生物量来满足其空间拓展和繁殖需求[12]。随着海拔的升高,可能由于水、气、光、热组合发生变异,气候条件变差,风力变强,有效积温降低,植物为了生存繁殖,调整生存对策改变细根生物量来适应严酷的高海拔环境。高海拔地区土壤酶活性降低[13],微生物代谢变慢抑制了对根系凋落物的分解速率,且海拔高的地方死根生物量相对较高,以此来维持局部小环境的温度,来保证活根的生长,这可能也是高海拔地区植物适应环境的一种策略[14]。还可能因为青藏高原是全球气候变暖的“启动器”和“放大器”,其温度升高要快于全球平均值[15]。海拔越高的地区气候变暖趋势越明显,高海拔地区森林环境因子向低海拔地区趋近[16],导致高海拔区域细根生长相对较快。另外,本研究仅选择海拔2175—3852 m细根生物量来研究分析,海拔继续升高后是否还有同样的规律有待进一步研究考证。
差异显著性分析表明5种林分间细根生物量差异不显著,林分和土层以及林分与海拔梯度的交互作用(表4)均对细根生物量没有显著影响,说明林分的变化并不影响细根生物量空间上的生态位[17]。热带落叶阔叶林的细根生物量为40.68 t/hm2、暖温带落叶阔叶林的为13.71 t/hm2、寒温带落叶阔叶林的为6.55 t/hm2和寒温带常绿针叶林的为7.31 t/hm2[18],可见细根生物量随着纬度的升高而减小。青海省森林细根生物量平均为8.50 t/hm2(所有样地细根生物量取均值),除山杨林,其他两种阔叶林的细根生物量比针叶林的细根生物量低,这与一般阔叶林细根生物量高于针叶林的有关研究结论不一致[19],因为植被群落类型不同,它们的生活对策不同,对光合作用产生的能量分配也不同,进而导致细根生物量出现差异。何永涛的调查发现白桦林细根生物量为7.89 t/hm2[20],与本研究结果相近。郭忠玲等研究表明[21],白桦林细根生物量最高(5.13 t/hm2)、其次为山杨林(3.94 t/hm2),这比本研究中的白桦林细根生物量低33.8%,比山杨林的低55.9%,可能是因为本研究区域所处的气候带[22]、海拔等地理因素不同导致土壤类型、立地条件有差异[23],也可能是林下草本群落组成尤其是优势种的不同引起的差异。
3.2 土壤理化性质的变化
土壤作为植被赖以生存的载体,理化性质在时间和空间上是异质性分布的[24]。低海拔区域降水量适宜,它可以明显影响土壤的导电率和含水率,致使土壤吸水溶胀,因而低海拔处土壤容重变小[25]。随着海拔的升高容重变大,可能由于海拔的变化导致林分改变,林分的变化又与土壤容重密切相关[26],林分可改变土壤的物理性状。还可能是海拔的变化引起土壤结构、质地、有机质含量差异等因素造成的。
土壤TC随海拔梯度先增加后减少,由于低海拔处气温较高,微生物以及酶活性较高,有机碳分解快土壤有机碳积累变少,区域含碳量以无机碳为主从而TC含量较少[27]。随着海拔的增加,温度降低,林地植被覆盖度高,地下地上生物量充足,微生物以及根系活性较弱,碳酸钙沉淀减少,有机质积累变多最终促使TC含量增加[28]。当海拔进一步升高,气温降低到一定界值,植被生存环境恶劣,作为TC重要来源的地上生物量相比其他区域较少,因而土壤TC含量降低。土壤TN含量随海拔的升高先增加后降低,可能是不同海拔林地植被凋落物数量和种类以及林地温度、湿度差异造成的[29]。土壤TN含量还可能受植物生长所消耗以及有机质分解氮的影响,二者相互作用导致TN随海拔先增后降[30]。土壤TP随海拔梯度逐步减少,可能因为P元素主要来源于岩石风化,土壤中P的迁移转化以及磷沉降容易受外界因子的影响而发生变异。海拔高度的增加,气候变化更明显,气候又是影响土壤P空间分布的主要因素[31],所以土壤TP随海拔的增加空间分布不规律。
各林分类型之间土壤容重均有差异,可能由于不同林分林下土壤母质、凋落物特性、根系分布存在差异导致的[32]。云杉、白桦林土壤容重较小,原因可能是:林地土壤团粒较多土壤疏松,从而土壤孔隙度大,容重变小;地表腐殖质层厚度适中、凋落物产量适宜,土壤微生物活性良好有机质分解较快,使得林地土壤疏松多孔,透气性好。所有林分表层容重显著小于深层,这和林地表层土壤中较高的资源有效性、微生物活性以及水肥耦合性有关[33]。
林分间土壤TC含量有差异,可能因为不同林分的树种组成有差异,植被冠层的光合固碳能力不同,导致林地凋落物的类别、质量和产量有差异,最终影响森林土壤的碳汇功能[34]。林分间土壤TN含量存在差异可能因为不同林分土壤的硝化速率不同,影响林分土壤氮素的转化,导致林分土壤的氮素水平产生差异。不同林分所处的地理位置、气候、土壤母质以及地表凋落物不同,又因为土壤中的P不只来源于凋落物的分解,还和土壤母质的矿物成分有关,所以林分间土壤TP含量存在差异。此外,云杉林的TC、TN、TP含量较高充分说明,作为青海省特有的优势树种,云杉对青海省森林土壤环境的优化和土壤生态平衡的维持意义重大[35]。
3.3 细根生物量与环境因子的相互关系分析
细根生物量与环境因子的相互影响关系已经有诸多报道。例如McCormack等[36]认为细根生物量与气候条件等因素有关。本文SEM显示土层、土壤容重直接影响细根生物量。土层是影响细根生物量的重要因素,细根生物量随着土层深度的增加而减少[37],且有大概70%集中在0—20 cm土层[38]。这是由于随着土层深度的增加土壤养分含量减少,影响细根吸收养分,还引起土壤下层温度变低细根的呼吸速率变小,致使细根生物量垂直下降[39]。土壤容重影响细根生物量,主要和土壤中的空气含量有关[40]。表层土壤容重较小,根系生长空间大且容易获得足够的空气,深层土壤空气含量减少,导致细根生物量减少[41]。由于不同林分地表凋落物的构成及其储量和分解速率有差异,造成林分间土壤容重不同[42],林分通过影响土壤容重间接影响细根生物量。
土壤的C、N、P元素是养分循环和转化的核心,它在驱动和调节森林生态系统演替过程发挥重要作用[43]。本研究发现细根生物量对土壤TC、TN、TP有较大影响。因为细根在森林生态系统凋落物和养分循环中,且作为根系中最为活跃的部分,它的不断生长、死亡以及分解变化对补偿土壤养分发挥着重要作用[44]。细根是C进入土壤生态系统的主要路径,大部分土壤C可能来源于它[45],细根死亡分解不仅对土壤N的贡献率比凋落物大18%—58%,而且还是归还土壤P的重要途径,倘若忽略细根的生产、死亡和分解作用,土壤养分的周转将会被低估20%—80%[46],所以细根生物量显著影响土壤养分。
4 结论
本研究分析了青海省森林细根生物量及其影响因子,表明青海省0—40 cm土层森林细根生物量平均为8.50 t/hm2,并且对土层、土壤容重和海拔的变化十分敏感。林分可能通过土壤容重间接影响细根生物量。同时,细根生物量还直接显著影响土壤养分含量。
