雅江流域干热河谷不同植被类型对土壤可蚀性的影响
2019-07-03姚甜甜
张 鹏 姚甜甜 喻 武 万 丹 叶 红
( 1. 西藏农牧学院资源与环境学院,西藏 林芝 860000;2. 西藏农牧学院高寒水土保持研究中心,西藏 林芝 860000)
土壤侵蚀是全球性的生态环境问题之一,土壤侵蚀现象的持续发生不仅使土壤质量不断下降,而且还会引起水体环境恶化等一系列生态问题[1-3]。影响土壤侵蚀的因素除了降雨、地形地貌、地表植被、人类活动等外在因素外,还与土壤自身性质和结构等内在因素密切相关[4],国际上通常使用可蚀性K值这一指标来衡量[5]。目前K值的计算方法主要有美国通用土壤流失方程(USLE)[6-9]、WEPP[10-11]和 EPIC[12]等 模 型[13],这些模型早期根据土壤属性计算土壤可蚀性K值,后期被诸多学者应用到林地、草地等土地利用方式[14-16]。目前,EPIC模型广泛应用于黄土高原、华北和东北等地区,对土壤侵蚀敏感性评价、土壤流失量预测等方面提供重要依据[17]。已有部分学者[5,18-19]采用EPIC模型对西藏自治区的土壤进行可蚀性K值进行计算,这对青藏高原的土壤可蚀性研究与应用具有重要的指导意义。
雅鲁藏布江中下游干热河谷区,夏季雨水充沛,山高坡陡,植被稀少,土壤结构破碎,极易产生水土流失。干热河谷中不同的植被类型通过改变植物根际、地上和地下枯枝落叶层,影响截留和下渗过程从而影响有机质的数量、质量及机械组成等因素,使得土壤物理性质发生显著变化,进而引起土壤可蚀性K值的变化[20]。近年来,随着西藏经济快速的发展,雅江流域的开发建设越来越剧烈。在桥梁、大坝等工程建设和人类不合理的放牧、樵采等活动影响下,雅江流域的生态环境遭受了一定的破坏,产生大量的水土流失,如何保护好雅江流域的生态环境是当前面临的重大问题。目前有关雅江流域干热河谷的研究主要集中在地质构造演化与植物分布等方面[21-23],而植被类型对土壤可蚀性的影响鲜有报道。因此,本研究选取雅江流域干热河谷4种典型植被——巨柏(Cupressus gigantea)、高山松(Pinus densata)、砂生槐(Sophora moorcroftiana)、铁杆蒿(Heteropappus altaicus),以其群落下的土壤为研究对象,对土壤可蚀性及其影响因素进行分析,旨在为区域水土流失防治提供参考。
1 研究区概况
研究区域主要位于西藏朗县境内,地处东经92°58′42.54 ″~93°36′09.78 ″,北纬 28°59′46.32″~29°09′57.00″,属雅鲁藏布江中下游干热河谷地带,喜马拉雅山北麓,平均海拔3 100 m。该区属温带大陆性半湿润气候,年平均气温11.0 ℃,年平均最高气温19.1 ℃,年平均最低气温5.3 ℃,无霜期130~170 d,年日照时数2 000~2 500 h,年均降水量350~600 mm。受印度洋暖湿气流西南季风的影响,6—9月为雨季,多为夜雨,其降雨量占年降雨量的90%以上。受印度洋暖湿气流东北季风影响,10月至次年5月为旱季,气候晴朗且干燥。朗县植物种类稀少,主要植被有高山松、巨柏、砂生槐、川滇高山栎(Quercus aquifolioides)、架棚(Ceratostigma minus)、忍冬(Lonicera japonica)、铁杆蒿等。
2 研究方法
2.1 样品采集与处理
于2017年9月,沿306省道,在研究区选取高山松群落、巨柏群落、砂生槐群落、铁杆蒿群落4种典型植被类型。每个植被类型内选取具有代表性的位置,设立5个10 m×10 m样方对样地植物多样性和生境特征进行调查(表1),并按照五点采样法,取0~20 cm土层原状土壤5个,用以测定土壤团聚结构、颗粒组成及有机碳。
每个样地同时按照五点采样法采取环刀土样10个,用于测定土壤容重、孔隙度、持水性等物理指标。采回原状土样按照纹路掰成直径为10 mm左右的团聚体,并将同一样方5个土样均匀混合进行自然风干。分取风干土样100~200 g 3份,进行团聚体分析。过0.149 mm孔径筛的土壤,用于有机碳的测定。
2.2 测定方法
土壤机械组成稳定性团聚体及水稳性团聚体采用沙维诺夫法[24]测定,土壤有机碳含量采用浓硫酸-重铬酸钾外加热法测定,土壤容重、孔隙度采用环刀法测定[24]。土壤机械组成采用Bettersize 2 000激光粒度分布仪分析,按照美国制设置颗粒分布;土壤pH采用美国IQ150土壤pH计测定。
2.3 指标值计算方法
采用Williams等[25]提出的EPIC模型计算土壤可蚀性K值,公式为:
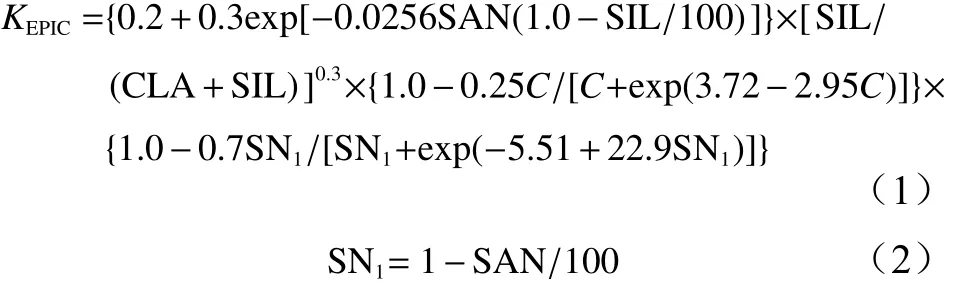
式中:SAN[26]表示砂粒(0.05~2.0 mm)含量(%),SIL表示粉粒(0.002~0.05 mm)含量(%),CLA表示黏粒(<0.002 mm)含量(%),C表示有机碳含量(%)。

式中:PAD为团聚体破坏率,S1为粒径大于0.25 mm团聚体干筛分析值,S2为粒径大于0.25 mm团聚体湿筛分析值。
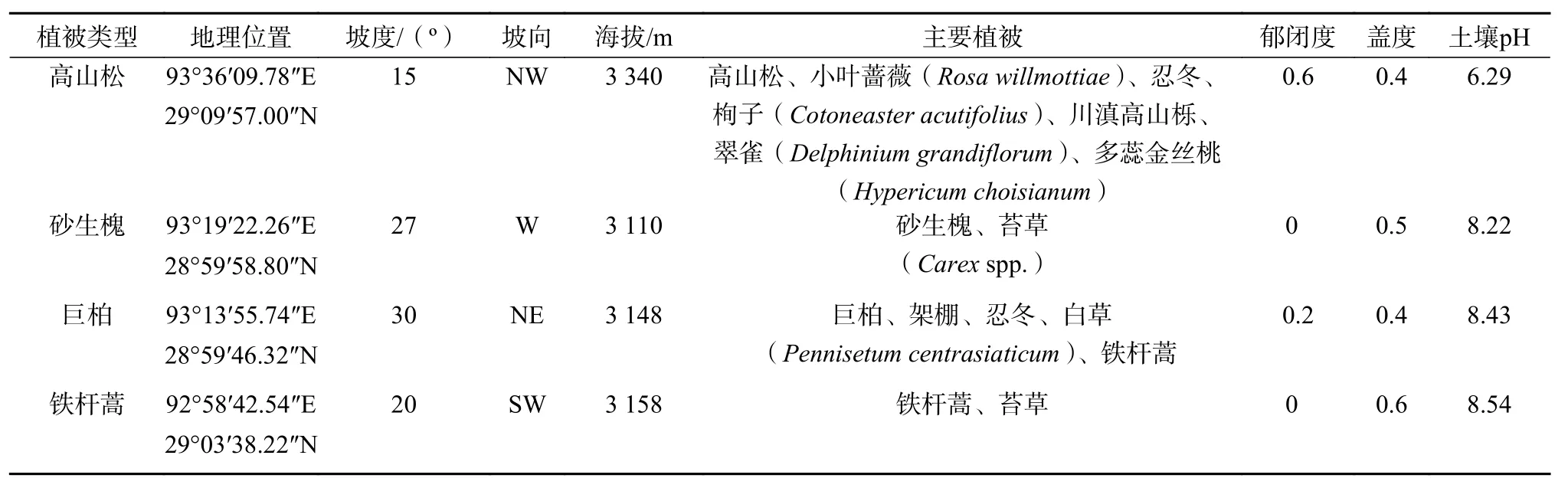
表 1 样地基本状况Table 1 Basic situation of samples
2.4 分析方法
数据统计、处理及图表分布采用Excel 2010、SPSS 17.0统计软件,不同数据组间差异显著性比较采用单因素方差分析(one-way ANOVA)和Duncan多重比较,相关性分析采用Pearson双变量相关分析。
3 结果与分析
3.1 土壤团聚体特征分析
由表2可知,4种植被类型下的土壤中,高山松的>0.25 mm的机械稳定性团聚体百分含量显著大于其他植被类型(P<0.05);在各粒级百分含量中以<0.25 mm的粒径所占的比例最大。其中在>0.25 mm的粒级中,高山松下土壤以>10 mm粒级所占比例最大,占20.13%,而1~2 mm粒级所占比例最小,为1.43%。砂生槐和巨柏下的土壤粒级团聚体均以0.5~1 mm和0.25~0.5 mm含量最高,分别为18.49%、20.47%和15.77%、22.09%,偏细粒化,而>2 mm的粒级团聚体以砾石为主。铁杆蒿在>0.25 mm的粒级中,以>10 mm粒级含量最高,为15.44%,3~5 mm粒级次之,为6.19%,7~10、5~7 mm 2个粒级分别为3.84%、3.04%,而0.5~1、0.25~0.5 mm 2个粒级团聚体总含量仅占到9.16%。综上所述,研究区不同植被类型下的土壤中,以<0.25 mm粒级所占比例最大,而在>0.25 mm粒级中,高山松和铁杆蒿集中在较大粒级,砂生槐和巨柏主要集中在较小粒级。
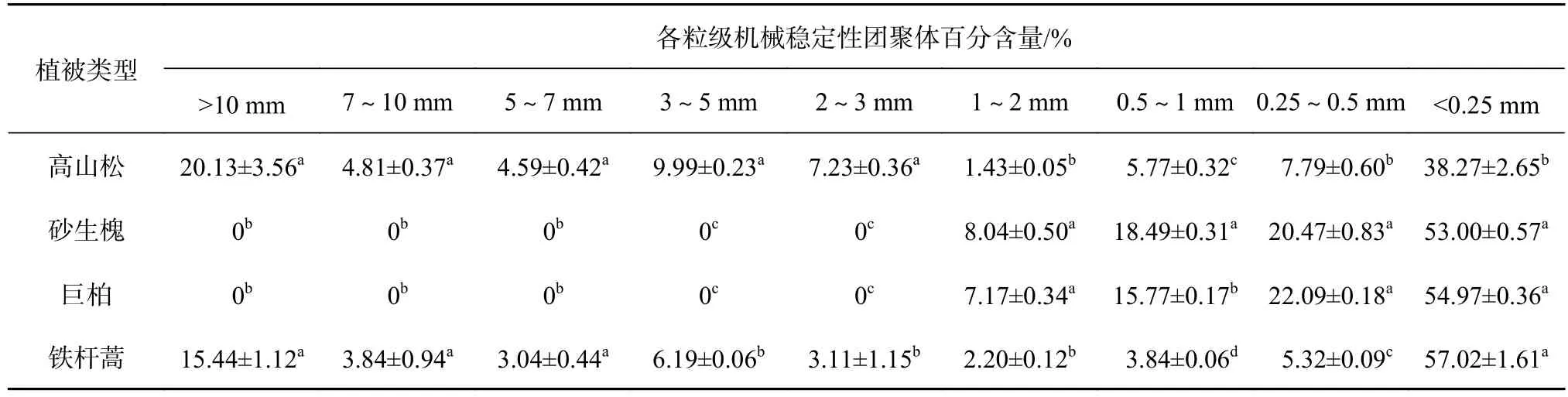
表 2 土壤机械稳定性团聚体分布特征Table 2 Distribution characteristics of soil mechanical stable aggregates
由表3可知,4种植被类型下的土壤>0.25 mm水稳性团聚体均占较高比例,在75.05%~82.97%,依次为巨柏>铁杆蒿>砂生槐>高山松。高山松以>5 mm粒级最大,达36.46%,2~5 mm粒级次之,为14.10%。砂生槐和巨柏主要集中在0.5~1 mm粒级,分别为42.25%、38.65%,0.25~0.5 mm团聚体次之,分别为22.17%、29.04%。铁杆蒿以>5 mm粒级最大,达43.11%,2~5 mm粒级次之,为19.05%,以0.25~0.5 mm粒级最小,为2.61%。总体而言,经湿筛后,研究区不同植被下的土壤团聚体仍能以>0.25 mm团聚体为主,其对以水力为主的外营力敏感度低,抵抗水流侵蚀能力较强。从团聚体破坏率看出,高山松和砂生槐较大,为24.95%和22.46%,铁杆蒿次之,为20.54%,巨柏最小,为17.03%。可见,高山松由于郁闭度高,枯落物覆盖较厚,释放有机酸等化学物质,使灌草盖度降低,浅层土壤受到灌草根系连结及改良作用变弱,导致土壤团聚体结构稳定性降低[27]。巨柏生长在坡陡的高山峡谷中,受人类干扰较小,使其土壤结构的稳定性最高。
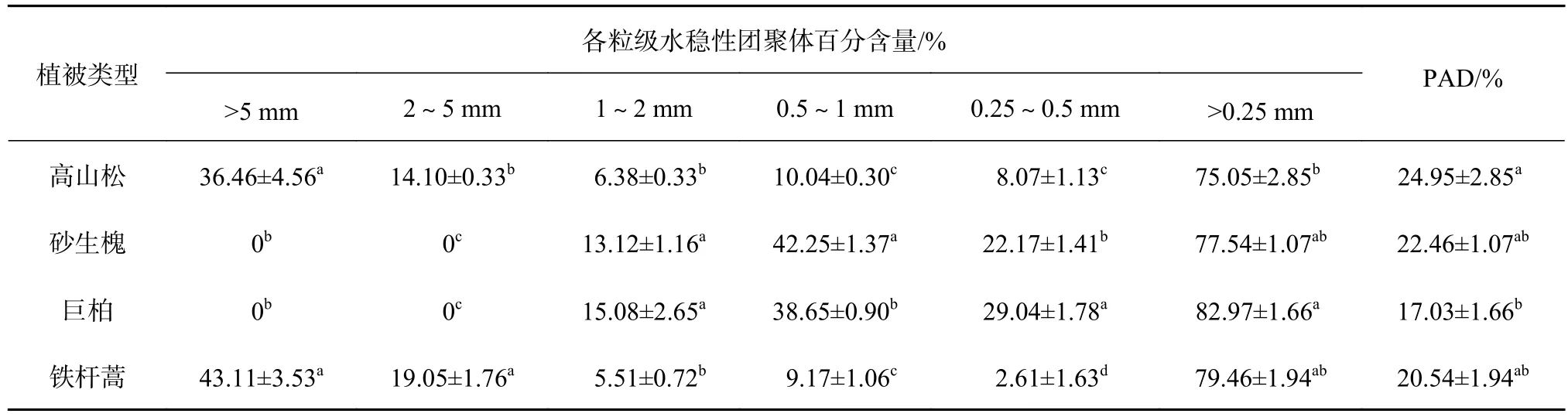
表 3 土壤水稳性团聚体及团聚体结构破坏率分布特征Table 3 Distribution characteristics of water stable aggregate and aggregate structure destruction rate
3.2 土壤颗粒组成及有机质分析
土壤粒径分布影响着土壤的肥力状况、水力特性及土壤侵蚀等,是重要的土壤物理特性[27]。由图1可知,4种植被类型下的土壤主要以粉粒、砂粒为主,黏粒含量仅占到0.22%~1.26%。按粒级组成划分均属砂质壤土,符合西藏高原地质历史年轻的特点。4种植被类型的土壤黏粒含量依次为:砂生槐>高山松>铁杆蒿>巨柏;粉粒含量依次为:高山松>砂生槐>铁杆蒿>巨柏;砂粒含量依次为:巨柏>铁杆蒿>高山松>砂生槐。不同植被类型间的土壤砂粒含量差异显著,粉粒含量高山松与砂生槐差异不显著,黏粒含量高山松与铁杆蒿两者差异不显著。砂粒含量中,4种植被类型土壤均大于70%。综上所述,研究区在雨季的降水冲刷下,水土流失严重,粗颗粒在水流的冲刷、搬运与沉积过程中,使其细小颗粒增多。土壤有机质可以促进团聚体的形成并保持其稳定性,提供微生物生命活动所需的能量[28]。4种植被类型的土壤有机质含量在20.19~37.99 g/kg之间,土壤肥力整体偏低。由图2可知,砂生槐、巨柏与铁杆蒿下的土壤有机质差异不显著,其高山松下的有机质含量显著高于其他3种植被(P<0.05)。这种理化性质的差异主要是因为研究区位于干热河谷,不同的海拔梯度使其水分产生差异,进而形成了不同的植被类型,其土壤空间变异性大,最终导致土壤物理性质产生分化。
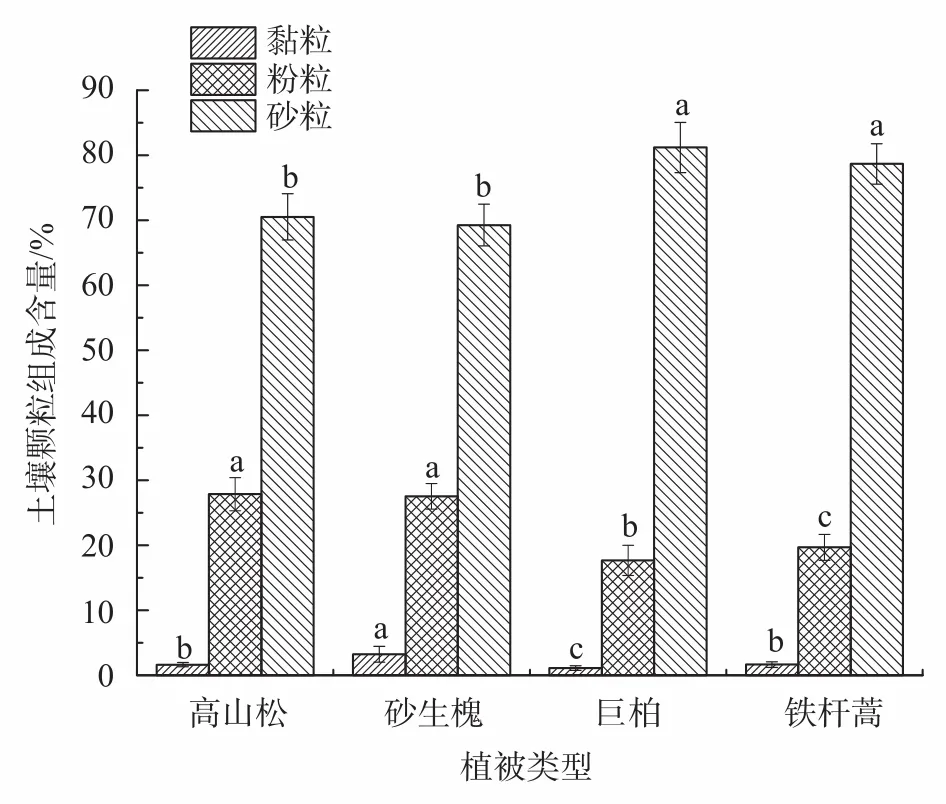
图 1 土壤颗粒组成分布特征Fig. 1 Distribution characteristics of soil particle size composition
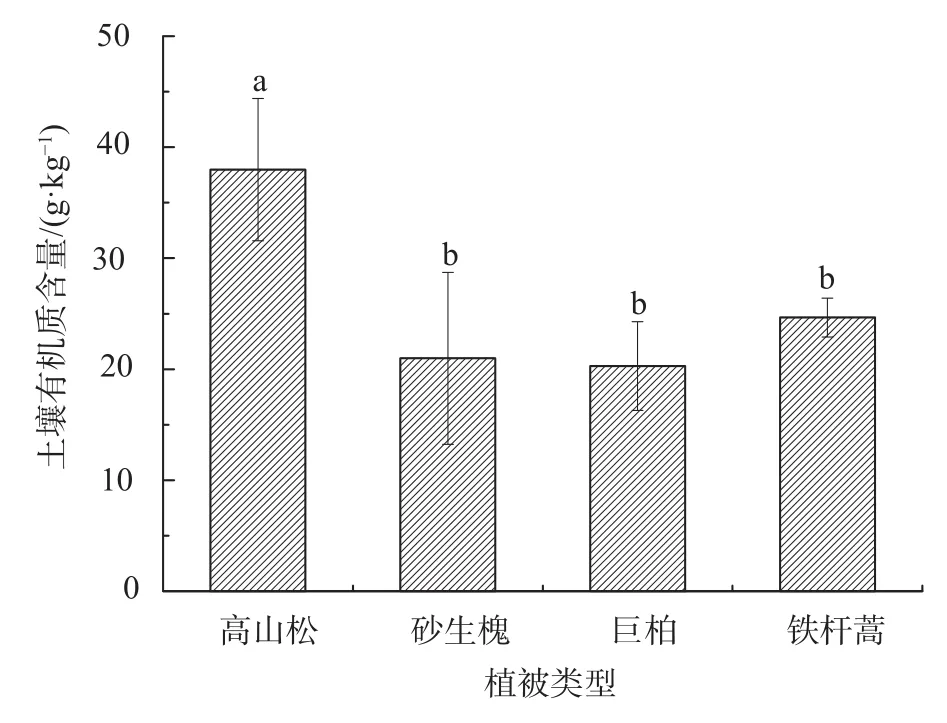
图 2 土壤有机质含量分布特征Fig. 2 Distribution characteristics of organic matter content
3.3 土壤容重及孔隙度分析
容重值能综合反映土壤结构、松紧状况、孔隙状况和土壤内生物的活动[29],土壤孔隙度能反映土壤的蓄水和通气性能。土壤容重越小,孔隙度越大,说明土壤结构越好,越有利于水分的保持与渗透[30-31]。由表4可知,不同的植被类型下,土壤容重彼此间呈极显著差异(P<0.01),其大小规律依次表现为:砂生槐(1.38 g/cm3)>巨柏( 1.28 g/cm3) >铁 杆 蒿 ( 1.04 g/cm3) >高 山 松(0.77 g/cm3),砂生槐、巨柏、铁杆蒿的土壤容重分别比高山松土壤容重高了79.90%、66.45%、36.29%。可见,这4种植被下的土壤中,高山松的土壤结构最好,保水保肥能力最强。这是由于高山松下的土壤枯枝落叶层覆盖厚,土壤的有机质含量高,促进了微生物间的活动,致使土壤的孔隙度增大,加之生长环境所需水分多,海拔相对较高,不易受人类的干扰,减少了对土壤的踩踏,使土壤的紧密程度低。
从土壤孔隙度来看,土壤的总孔隙度均值由大到小依次表现为:铁杆蒿(70.05%)>高山松(66.57%)>巨柏(58.94%)>砂生槐(51.84%),高山松土壤总孔隙度与铁杆蒿差异不明显,与其他植被类型差异均显著(P<0.05)。各植被间毛管孔隙度的差异性变化与总孔隙度的一样,非毛管孔隙度差异均不明显。总体来看,高山松和铁杆蒿土壤的总孔隙度和毛管孔隙度明显高于其他植被类型。土壤非毛管孔隙数量大小能体现土壤渗透和滞留水分能力强弱[32],一般非毛管孔隙度大于10%,能够有效地改善土壤结构[27]。分析4种植被类型的土壤,非毛管孔隙度都小于10%。一般认为,结构性良好、通气性强的土壤,孔隙度分布在40%~50%之间,非毛管孔隙度大于10%,而非毛管孔隙度与毛管孔隙度比例在1∶4~1∶2[33-34]。由表4可知,4种植被类型的土壤均不在良好比例范围之内。由此说明,雅江流域干热河谷地带的土壤与肥沃的土壤相比较,通气性、透水性与涵养水源能力均较差,水土流失中携带土壤能力强,一旦现存仅有的地表植被被破坏,所产生的水土流失规模更大。
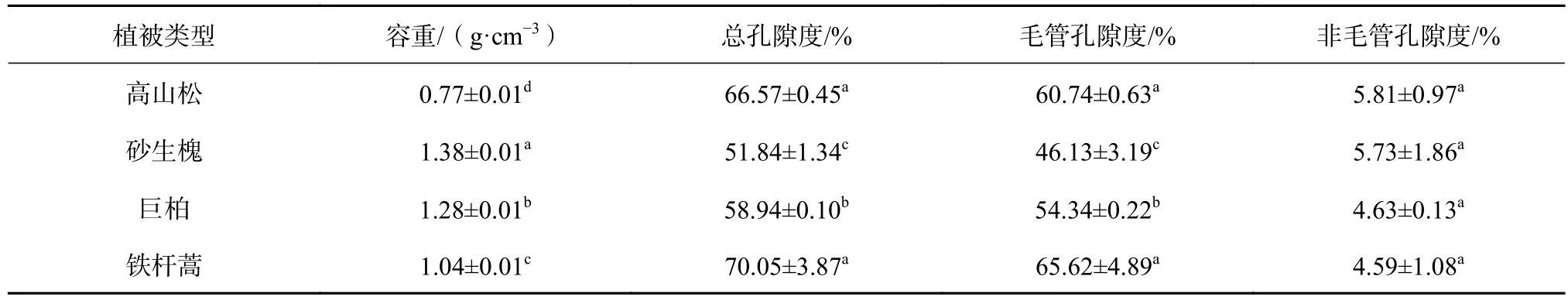
表 4 不同植被类型下的土壤物理特性Table 4 Physical characteristics of soil in different vegetation types
3.4 土壤贮水性能分析
土壤含水量是表明土壤水分状况的一个指标,对土壤形成发育过程及肥力水平高低等有重要影响[27]。由表5可知,不同植被类型土壤自然含水率依次为:高山松(38.51%)>铁杆蒿(27.68%)>巨柏(17.63%)>砂生槐(14.13%)。砂生槐自然含水率与巨柏差异不明显,与其他植被类型均差异显著(P<0.05)。这说明砂生槐和巨柏生长环境所需土壤的水分远小于铁杆蒿和高山松。高山松作为乔木,其生长环境所需要的水分远远高于砂生槐根系的水分,致使高山松的土壤自然含水率最高,砂生槐最低。
对不同植被类型下土壤贮水能力进行方差分析。由表5可知,砂生槐与巨柏无显著差异,高山松与铁杆蒿无显著差异。这主要是因为砂生槐和巨柏的土壤偏砂性,细小的粒级含量多,土壤贮水能力差。高山松与铁杆蒿土壤虽然也偏砂性,但是较大的粒级含量高于砂生槐和巨柏,土壤的贮水能力也相应增加。4种植被的毛管持水量与饱和持水量差异显著(P<0.05)。砂生槐的土壤饱和持水量与土壤贮水能力均显著小于其他植被类型,分别为376.23 t/hm2和38.99 t/hm2,高山松的土壤饱和持水量与毛管持水量均最高,分别为868.81 t/hm2和792.65 t/hm2。这说明在山高坡陡的雅江河谷,高山松生长环境的山体海拔高于生长于河谷边的砂生槐,在雨季的冲刷下,高山松下高海拔土壤中的细颗粒被冲刷至低海拔,致使高山松的土壤粗颗粒远远大于砂生槐,土壤持水能力也远远高于砂质土壤下的砂生槐。

表 5 不同植被类型下的土壤持水能力Table 5 Soil water-holding capacity in different vegetation types
3.5 不同植被类型土壤可蚀性分析
土壤可蚀性是评价土壤是否易受侵蚀营力破坏的性能,也是土壤对侵蚀营力分离和搬运作用的敏感性[35]。采用EPIC经验公式,对不同植被类型下土壤的可蚀性K值进行计算,结果详见表6。不同植被类型下土壤可蚀性K值由大到小依次为:砂生槐>高山松>巨柏>铁杆蒿。研究区土壤可蚀性K值分布范围在0.152 2~0.246 0,4种植被下的土壤可蚀性K值均值为0.183 2,其中砂生槐土壤可蚀性K值最大,为0.223 0,铁杆蒿的土壤可蚀性K值最小,为0.156 3。根据刘斌涛等[5]对青藏高原的土壤可蚀性强弱的分级标准,除了砂生槐土壤为中等可蚀性外,其余三者都是较低可蚀性。CV值介于0.01%~0.60%之间,说明研究区同一种植被土壤可蚀性K值的空间变异性较弱。4种植被类型土壤可蚀性K值的差异性显著(P<0.05),这说明不同植被类型受成土条件影响,土壤抗侵蚀能力差异显著[19]。原因是研究区不同植被下的土壤受其地形、海拔梯度、水分等小环境的影响,使其土壤理化性质产生差异。而土壤可蚀性是其内在理化性质决定的特征参数,K值确定性表现在其仅为土壤理化性质的参数[36]。K值的大小表示土壤抗侵蚀能力的强弱,K值越大,抗侵蚀能力越弱;反之,K值越小,抗侵蚀能力越强[37]。由此可知,相较于其他植被类型,砂生槐土壤抗侵蚀能力最差。这是因为砂生槐生长于干旱的河谷地带,加之其建群种单一,优势度高,致使灌草覆盖度减少,土壤结构稳定性变差。
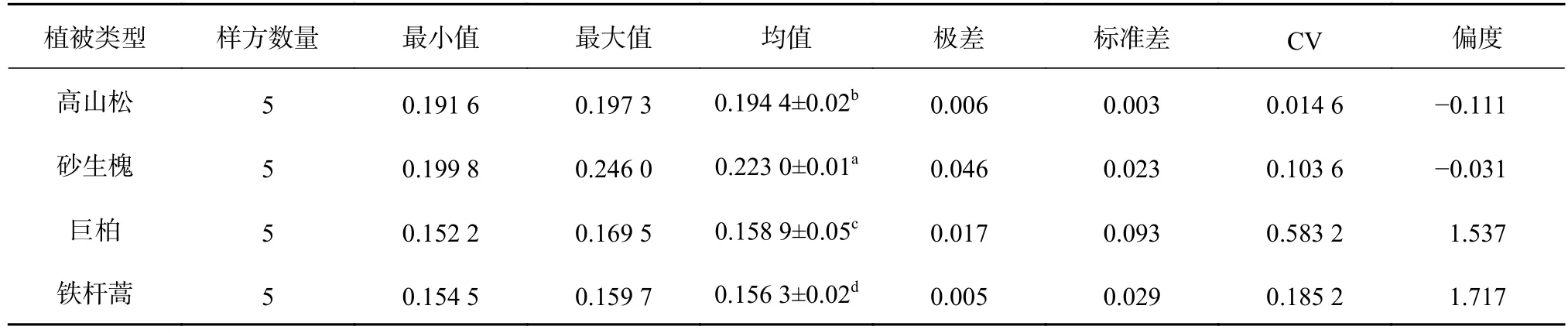
表 6 土壤可蚀性K值统计特征Table 6 Statistical characteristics of K value of soil erodibility
3.6 土壤可蚀性与影响因子相关性分析
由于土壤可蚀性仅受其自身理化性质的影响而发生相应变化[36],EPIC模型仅反映出所估算的土壤可蚀性K值与机械组成、有机碳具有一定的相关关系,并未能反映其与土壤自身理化性质的相关程度。因此为进一步揭示土壤可蚀性受其自身物理指标的影响规律,对土壤的可蚀性K值与土壤机械组成、有机质含量、团聚体状况、土壤容重、孔隙度、土壤贮水能力、自然含水率进行Pearson相关性分析,结果见表7。土壤可蚀性K值与黏粒、粉粒含量呈极显著正相关关系,与砂粒含量呈极显著负相关关系,与总孔隙度、毛管孔隙度呈显著负相关关系,与非毛管孔隙度呈显著正相关关系。这是因为研究区土壤年轻,风化程度低,砂性很强,有大量的砾石所致。土壤可蚀性K值与土壤容重呈正相关,与总孔隙度呈显著负相关,这说明较为紧密的土壤能够有效减少土壤的渗透能力,增大地表的径流,从而增大了土壤的侵蚀能力[19]。从相关性分析来看,研究区不同植被的土壤可蚀性K值主要受到土壤颗粒机械组成以及孔隙度的影响。
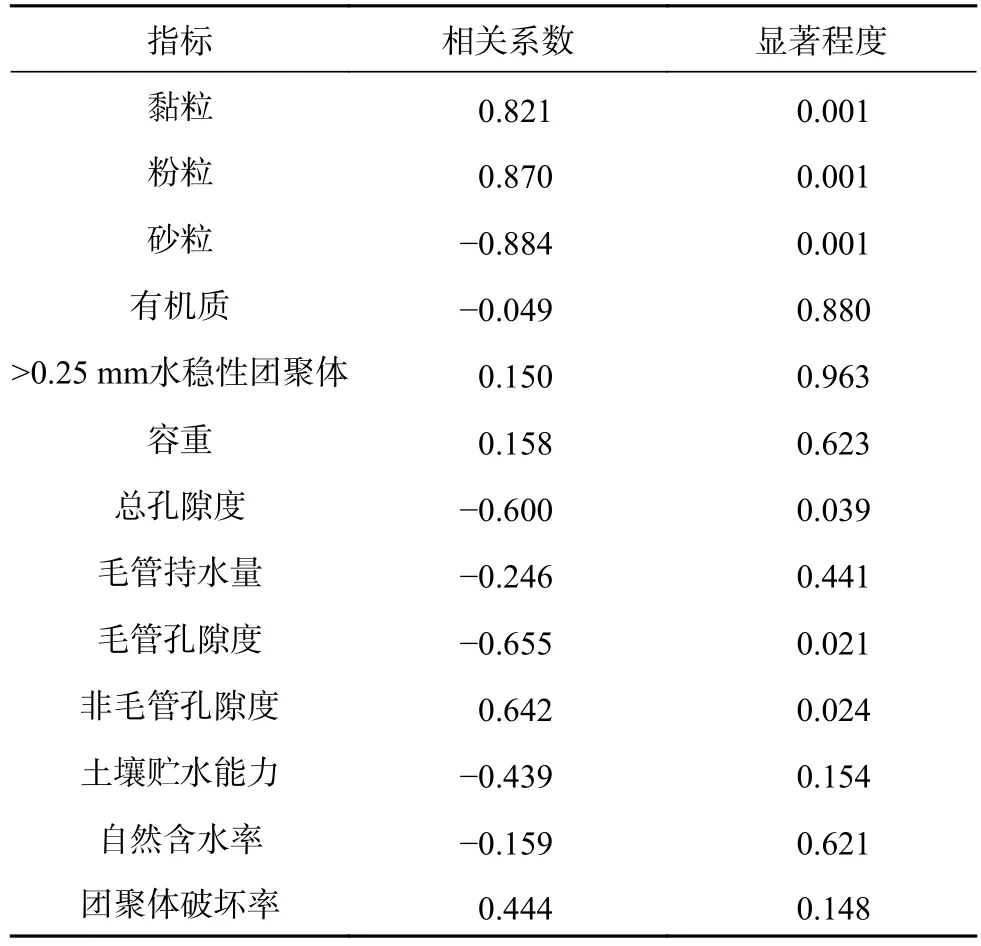
表 7 EPIC模型指标与可蚀性K值的相关性Table 7 Correlation between EPIC model index and K value of erodibility
4 结论与讨论
4.1 结论
1)研究区4种植被下土壤容重差异显著,砂生槐的土壤总孔隙度显著小于高山松、巨柏与铁杆蒿的,铁杆蒿与高山松、巨柏的土壤自然含水率显著大于砂生槐的,砂生槐的土壤持水能力显著小于高山松、巨柏与铁杆蒿。
2)研究区土壤机械稳定性团聚体均以<0.25 mm所占的比例最大,经湿筛后,团聚体以>0.25 mm为主。研究区土壤主要以粉粒、砂粒含量为主,土壤质地偏砂性。土壤有机质含量在20.19 ~37.99 g/kg,肥力偏低。
3)土壤可蚀性K值分布范围在0.152 2~0.246 0,平均值为0.183 2,为较低可蚀性。土壤可蚀性K值由大到小依次为:砂生槐>高山松>巨柏>铁杆蒿。
4)土壤可蚀性K值与砂粒含量为极显著负相关,与粉粒和黏粒呈极显著正相关,与总孔隙度、毛管孔隙度呈显著负相关,与非毛管孔隙度呈显著正相关。
4.2 讨论
4.2.1 不同植被类型对团聚体的影响
水稳性团聚体是评价土壤结构稳定性与抗蚀能力的重要指标[38]。本研究显示,4种植被下的土壤机械稳定性团聚体>0.25 mm的粒级中,巨柏所占比例最小,高山松所占比例最大。经湿筛后,>0.25 mm水稳性团聚体中高山松所占比例最小,而巨柏所占比例最大,这说明高山松的土壤所含粗颗粒多,土壤相对呈原始状态,巨柏土壤偏小粒径级,这也符合干热河谷气候下高山松生长在山顶地带,巨柏生长在坡下位置的特点。高山松在山体坡上位置土壤呈相对原始状态,在水流的作用下,粗颗粒含量高的高山松的土壤冲刷一部分成细颗粒,细颗粒被冲刷至下坡位置,使得土壤的抗侵蚀能力小于偏砂性的巨柏。
4.2.2 不同植被类型对土壤可蚀性K值及相关因子的影响
研究区4种植被下的土壤有机质含量在20.19~37.99 g/kg之间,土壤肥力较低,这是因为研究区的地质历史年轻,土壤以粉砂粒含量为主,而有机质主要在黏粉粒中,致使土壤的有机质含量低。而在有机质含量中高山松最大,铁杆蒿次之,巨柏和砂生槐的含量均最小,这是由于受雅江干热河谷气候的影响,高山松的生长环境位于高山的上部,砂生槐和巨柏生长于雅江河谷底部,致使高山松的水分相对较多,植被新陈代谢旺盛,枯枝物生物量也较多,使高山松的土壤有机质含量最高。研究区4种植被下的土壤可蚀性K值范围是0.152 2~0.246 0,平均值为0.183 2,较刘斌涛等[5]得出的藏南阿嘎土和寒钙土地带的土壤可蚀性平均值低,属较低可蚀性。前者是通过模型计算和面积加权的方法来定量研究西藏土壤种类的土壤可蚀性,而本研究细化了干热河谷区的土壤可蚀性。受水热条件的影响,不同植被类型下的土壤理化性质存在差异,进而影响了土壤的可蚀性。4种植被的土壤可蚀性K值由大到小依次为:砂生槐>高山松>巨柏>铁杆蒿。铁杆蒿为0.156 3,巨柏为0.158 9,高山松为0.194 4,砂生槐是0.223 0。可以看出,同属砂质土壤的砂生槐高于巨柏。因为,巨柏下的植物丰富度远高于砂生槐,使得从高海拔冲刷下的颗粒拦截能力增加,加之生长于山高坡陡的地带和人类对巨柏的保护,使其下的土壤可蚀性K值也较小。砂生槐生长于雅江干热河谷及山坡的沙丘上,对比其他植被生长环境,土壤水分更稀少,致使其下的草本植物生长稀疏,加之受人类干扰的影响大,牲畜踩踏土壤,土壤渗透性降低,产生地表径流的能力更强,土壤的可蚀性也最大。高山松的团聚体破坏率最大,但是土壤可蚀性小于砂生槐,这也与高山松的植物丰富度远大于相对单调的砂生槐有关。高山松群落下的植被丰富度高,土壤的有机质含量高增强了团聚体的稳定性,土壤可蚀性也小于砂生槐。铁杆蒿植被覆盖度和有机质含量较高,土壤总孔隙度大,渗透能力强,自身拦截水流能力强,土壤的可蚀性也最小。
4.2.3 土壤可蚀性K值对不同理化指标因子的响应
研究区土壤可蚀性K值与砂粒含量为极显著负相关,与粉粒呈极显著正相关,与黏粒呈极显著正相关,这与前人研究结果一致,即土壤可蚀性K值随粉粒或黏粒含量的增加而增大,随砂粒含量的增加而减小[39],这是因为砂粒含量的增加使土壤的孔隙度增大,增强了地表的水分下渗,减小了地表的径流强度,使土壤侵蚀能力降低。土壤可蚀性K值与总孔隙度、毛管孔隙度呈显著负相关,与非毛管孔隙度呈显著正相关,这主要是由于黏粒含量增加使土壤水分下渗能力降低,堵塞了土壤孔隙,致使土壤渗透能力变差,地表径流强度增加,使其土壤侵蚀能力增大。研究发现,研究区土壤可蚀性在考虑土壤的自身物理性质外,还应进一步研究砾石含量、微生物量等对土壤可蚀性的影响[40]。
