模拟氮沉降对天山云杉林土壤酶活性的季节性影响
2019-07-03李吉玫张毓涛
李吉玫 张毓涛
( 1. 新疆林业科学院森林生态研究所,新疆 乌鲁木齐 830063;2. 新疆天山森林生态系统国家定位观测研究站,新疆 乌鲁木齐 830063)
干旱区温带森林通常表现为氮缺乏型生态系统[1],然而在过去一个世纪中,受人类活动的干扰,通过大气干湿沉降进入陆地生态系统的含氮化合物大量增加,而且氮沉降量在未来几十年将继续增加,还呈现出全球化趋势[2-3]。氮沉降增加对森林生态系统物质循环和能量流动及转换有显著影响。土壤酶在物质循环和能量流动中起着决定性的作用,能敏感地预警土壤生态系统发生的微小变化,土壤酶活性反映土壤中进行各种生化过程的方向和强度,氮沉降通过改变土壤有机质积累及分解、土壤矿化过程等改变土壤酶活性变化。土壤水解酶(蔗糖酶和脲酶)及氧化酶(过氧化物酶和多酚氧化酶)能很好地表征土壤碳、氮的供应程度、土壤腐殖化程度及有机质的积累程度,对这几种土壤酶活性的研究有助于深入了解土壤微环境对人类活动干扰的响应[4-5]。通常认为,长期氮沉降对氮缺乏型的生态系统中与木质素降解等有关的多酚氧化酶活性有抑制作用,但在氮饱和型生态系统得出了相反的结论。氮沉降的增加除对与碳相关酶活性有影响外,对氮相关酶活性也有正面或负面的影响[5]。但由于土壤酶活性受多种复杂因素的影响,如土壤水分、凋落物基质等。因此,不同生态系统土壤酶对氮沉降的响应可能有差异[6-8]。尽管如此,在干旱区森林生态系统中,有关氮沉降增加如何影响土壤酶的变化仍知之甚少。
天山云杉(Picea schrenkiana) 林是新疆山地森林的主要组成部分,在干旱区涵养水源、保育土壤、保护生物多样性和固碳释氧等方面发挥着重要的生态功能。有研究表明天山中部氮沉降量约 为 6.82~ 3.85 kg/( hm2·a), 平 均 达 5.33 kg/(hm2·a)[9-10]。同时,天山中部天山云杉林分布的部分区域由于重大工程的施工,如煤矿开采、水泥厂的修建等造成该区域氮沉降量有逐渐增加的趋势。本研究以天山森林生态系统国家定位观测研究站为依托,通过2 a的野外模拟试验,研究氮沉降对天山云杉林林下土壤酶活性的影响,为了解氮沉降对生态系统变化的影响提供参考。
1 试验区概况
试验区设在天山森林生态系统国家定位观测研究站,地处天山山脉中段北坡,位于东经87°27′28.5″~87°28′47.7″,北纬 43°24′48.3″~43°26′17.9″,海拔 1 908~2 960 m。该地区属温带大陆性气候,年均气温2.0 ℃,年降水量400~600 mm,雨季集中在6—8月份,年蒸发量980~1 150 mm,年均相对湿度65%,干燥度1.4,无霜期89 d,大于等于10 ℃积温1 170.5 ℃。林下土壤为山地灰褐色森林土,腐殖质层较厚。该区植被类型是以天山云杉为主的温带针叶林,森林覆盖率达60%。
2 试验方法
2.1 样地设置
2015年秋季(9月底)在研究区设置3 m×3 m的固定样地12个,每个样地间设置1 m的缓冲带,并进行林分状况和土壤养分等调查,基本情况见表1。样地内主要地被植物有:天山羽衣草(Alchemilla tianshanica)、羊角芹(Aegopodium podagraria)、天蓝岩苣(Cicerbita azurea)、木地肤(Kochia prostrata)、羊茅(Festuca ovina)、粗根老鹤草(Geranium dahuricum)等。

表 1 各处理样地的林分和土壤养分基本情况Table 1 Basic conditions of forest stand and soil nutrients in each treatment plot
2.2 氮沉降增加模拟
结合已有学者对天山中部氮沉降监测的结果,本试验设计4种氮处理,分别为CK (对照,不施氮)、LN(低氮,5 kg/(hm2·a))、MN(中氮,10 kg/(hm2·a))、HN(高氮,15 kg/(hm2·a)),每种处理重复 3 次。将 NH4Cl溶于30 L水中,每月月初无雨时均匀喷洒,于2015年9月底、2016—2017年5月初、7月初和9月初将尿素与细沙充分搅拌并均匀地洒在各个样地中。
2.3 取样方法
2016—2017年各次施完肥后的第3~5天,在每个样地沿2条对角线取土样,即每个样地5个样品,同一样地同层土样均匀混合,共取6次样。在取样时将凋落物层去除,分0~10 cm和10~20 cm 2层,将土壤样品装入有冰袋的采样箱,带回实验室,在4 ℃下保存,进行土壤酶活性和土壤养分测定。
2.4 土壤养分和酶活性测定方法
土壤碱解氮、速效磷及速效钾分别采用碱解扩散法、浸提-钼锑抗比色法及火焰光度法测定[11]。土壤脲酶活性采用苯酚-次氯酸钠比色法测定,用每克土样每小时生成的NH3-N的数量表示脲酶活性;土壤多酚氧化酶与过氧化物酶活性采用邻苯三酚比色法测定,用每克土样每小时生成的紫色没食子素量表示酶活性;蔗糖酶用3,5-二硝基水杨酸比色法测定,用每克土样每小时生成的葡萄糖数表示蔗糖酶活性[12]。所有的土壤样品均送至中国科学院新疆生态与地理研究所中心实验室测定。
2.5 数据处理
各氮沉降处理间土壤酶活性及不同季节土壤酶活性的差异用单因素方差分析(one way ANOVA),用最小极差法(LSD)进行各个处理间的多重比较。所有的统计和作图采用Origin 7.5完成。
3 结果与分析
3.1 氮沉降增加对天山云杉林土壤脲酶活性的影响
不同氮沉降处理下土壤脲酶活性特征见表2。由表2可知,不同处理土壤脲酶活性表现为HN>MN>LN>CK,但无显著差异。与CK处理的0~10 cm土层相比,LN、MH和HN处理土壤脲酶活性提高了8.82%~35.29%;10~20 cm土层提高了20.83%~54.17%。说明土壤脲酶活性受到底物刺激,提高了土壤脲酶活性。

表 2 不同氮沉降处理下土壤脲酶活性特征Table 2 Characteristics of soil urease activity under different nitrogen deposition treatments mg/g
不同季节氮沉降处理对土壤脲酶活性的影响见图1。季节不同,氮沉降处理对不同土层土壤脲酶活性的影响有差异,其中春季氮沉降增加对0~10 cm和10~20 cm土层土壤脲酶活性均没有显著影响。但与CK相比,LN、MN和HN处理0~10 cm土层土壤脲酶活性分别增加了6.54%、13.88%和15.44%;10~20 cm土层脲酶活性分别增加了10.81%、17.95%和19.44%。夏季氮沉降增加后,0~10 cm土层除CK与LN差异显著外,其余处理土壤脲酶活性差异显著(P<0.05),但10~20 cm土层差异不显著。与CK相比,LN、MN和HN处理0~10 cm土层土壤酶活性分别增加了11.71%、16.44%和27.31%,10~20 cm增加了4.13%、6.93%和7.48%。秋季模拟氮沉降增加后,0~10 cm土层,除了MN和HN处理的差异不显著外,其余处理差异均显著(P<0.05),但10~20 cm土层各处理的脲酶活性差异均不显著。与CK相比,LN、MN和HN处理0~10 cm土层土壤酶活性分别增加了20.68%、36.06%和40.77%;10~20 cm土壤分别增加了5.30%、8.41%和8.86%。
3.2 氮沉降增加对天山云杉林土壤蔗糖酶活性的影响
不同氮沉降处理下土壤蔗糖酶活性特征见表3。由表3可知,随着氮添加量的增加,土壤蔗糖酶活性呈现先增加后降低的趋势,即MN>LN>HN>CK。但方差分析表明,氮添加仅对0~10 cm土层土壤蔗糖酶活性有显著影响,对10~20 cm土层的影响不显著。
不同季节氮沉降处理对土壤蔗糖酶活性的影响见图2。春季模拟氮沉降增加后,LN和MN处理0~10 cm土层蔗糖酶活性分别比CK增加了3.96%和 9.23%,HN处理降低了14.73%;10~20 cm土层分别增加了27.61%和49.45%,HN处理降低了16.67%。夏季模拟氮沉降增加后,LN和MN处理0~10 cm土层蔗糖酶活性分别比CK增加了14.59%和19.37%,HN处理降低了3.40%;10~20 cm土层分别增加了13.88%和19.21%,HN处理降低了13.40%。秋季模拟氮沉降增加后,LN和MN处理0~10 cm土层蔗糖酶活性分别比CK增加了7.38%和10.66%,HN处理降低了7.36%;10~20 cm土层分别增加了20.45%和29.79%,HN处理降低了11.79%。

图 1 不同季节氮沉降处理对土壤脲酶活性的影响Fig. 1 Effects of nitrogen deposition treatment on soil urease activity in different seasons
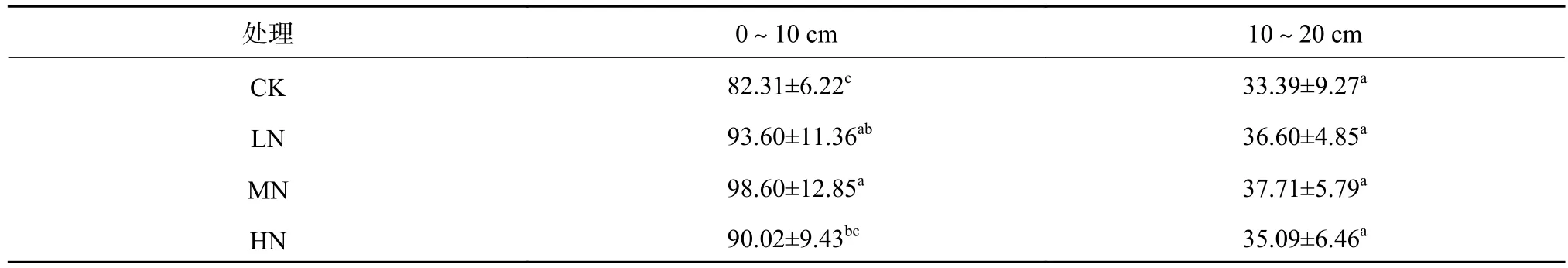
表 3 不同氮沉降处理下土壤蔗糖酶活性特征Table 3 Characteristics of soil sucrase activity under different nitrogen deposition treatments mg/g
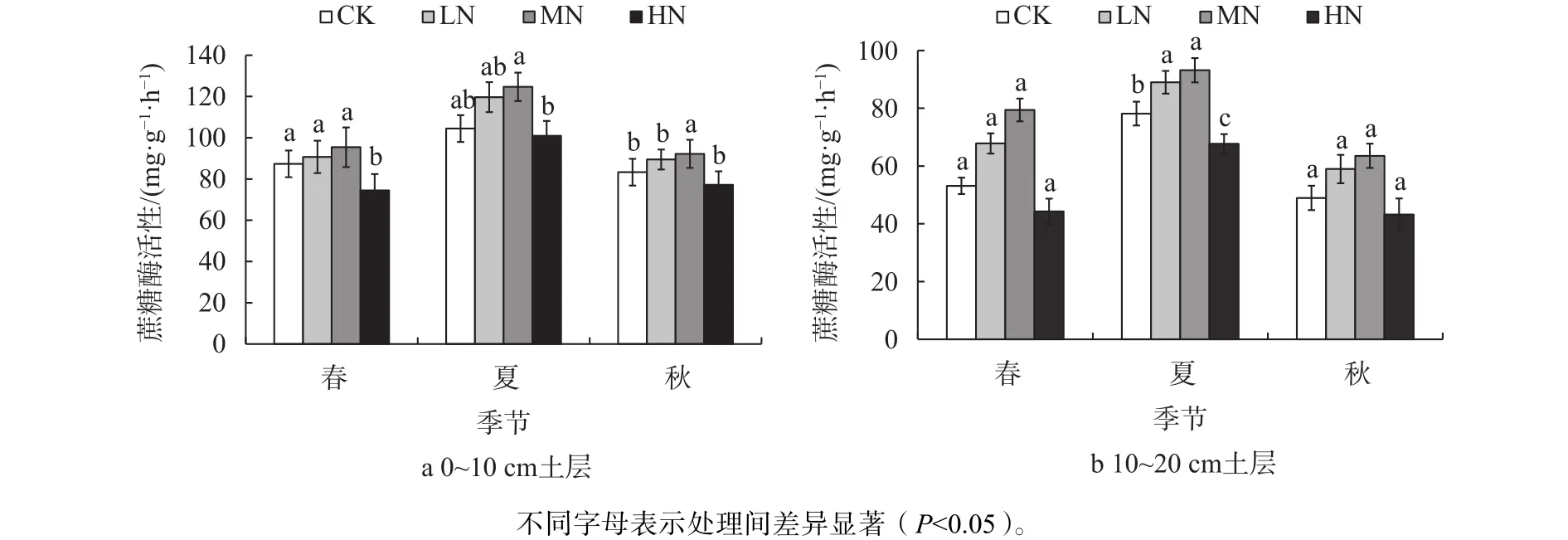
图 2 不同季节氮沉降处理对土壤蔗糖酶活性的影响Fig. 2 Effects of nitrogen deposition treatment on soil sucrase activity in different seasons
3.3 氮沉降增加对天山云杉林土壤过氧化物酶活性的影响
不同氮沉降处理下土壤过氧化物酶活性特征见表4。由表4可知,不同处理土壤过氧化物酶活性表现为CK>LN> MN> HN(表4)。且0~10 cm土层除了LN和MN处理间,差异不显著外,其余处理差异均显著(P<0.05);10~20 cm土层除了HN与其余的差异显著(P<0.05)外,其余处理间差异不显著。与CK处理的0~10 cm土层相比,LN、MH和HN处理土壤过氧化物酶活性降低了21.83%~37.21%;10~20 cm土层相比,降低了8.04%~31.98%。

表 4 不同氮沉降处理下土壤过氧化物酶活性特征Table 4 Characteristics of soil peroxidase activity under different nitrogen deposition treatments mg/g
不同季节氮沉降处理对土壤过氧化物酶活性的影响见图3。春季模拟氮沉降增加后,4种氮处理间0~10 cm和10~20 cm土层土壤过氧化物酶活性均没有显著差异(图3)。但与CK相比,LN、MN和HN处理0~10 cm土层土壤过氧化物酶活性分别降低了5.60%、16.79%和23.55%,10~20 cm土层分别降低了5.96%、19.58%和30.67%。夏季模拟氮沉降增加后,2层土壤过氧化物酶活性影响显著(P<0.05)。其中0~10 cm土层,CK和LN之间的差异不显著,但均与MN和HN差异显著(P<0.05);10~20 cm土层除了HN与其他处理的差异显著(P<0.05)外,其余处理间差异均不显著。与CK相比,LN、MN和HN处理0~10 cm土层土壤过氧化物酶活性分别降低了7.56%、20.21%和22.90%,10~20 cm降低了12.95%、21.34%%和28.46%。秋季模拟氮沉降增加后,0~10 cm土层土壤过氧化物酶活性差异显著(P<0.05),但对10~20 cm影响不显著。与CK相比,LN、MN和HN处理0~10 cm土层土壤过氧化物酶活性分别降低了23.45%、29.65%和57.37%,10~20 cm土层分别降低了4.25%、12.35%和42.61%。表明氮添加起到了抑制土壤过氧化物酶活性的作用,且随着土层的加深,抑制作用不断降低。

图 3 不同季节氮沉降处理对土壤过氧化物酶活性的影响Fig. 3 Effects of nitrogen deposition treatment on soil peroxidase activity in different seasons
3.4 氮沉降增加对天山云杉林土壤多酚氧化酶活性的影响
不同氮沉降处理下土壤多酚氧化酶活性特征见表5。由表5可知,不同处理间土壤多酚氧化酶活性的变化趋势与过氧化物酶活性的趋势基本一致,即表现为 CK>LN> MN> HN(表 5)。且方差分析表明氮沉降仅对其影响显著(P<0.05),说明氮添加对土壤多酚氧化酶活性起到了明显的抑制作用。与CK处理的0~10 cm土层相比,LN、MH和HN处理土壤多酚氧化酶活性降低了5.03%~23.19%;10~20 cm土层相比,降低了5.86%~14.52%。

处理 0~10 cm 10~20 cm CK 34.37±10.58a 27.83±8.76a LN 32.64±10.13a 26.20±8.74a MN 28.74±8.16b 25.20±8.35a HN 26.40±8.20b 23.79±8.38a
不同季节氮沉降处理对土壤多酚氧化酶活性的影响见图4。只有春季模拟氮沉降增加后,各处理土壤多酚氧化酶活性差异不显著(P<0.05)。春季模拟氮沉降增加后,LN、MN和HN处理0~10 cm土层多酚氧化酶活性分别比CK降低了7.66%、13.84%和19.17%;10~20 cm土层分别降低了6.14%、11.80%和13.92%%。夏季模拟氮沉降增加后,LN、MN和HN处理0~10 cm土层多酚氧化酶活性分别比CK降低了5.73%、12.27%和24.11%,10~20 cm土层降低了18.91%、22.24%和33.94%。秋季模拟氮沉降增加后,LN、MN和 HN处理 0~10 cm土层分别比 CK降低了10.98%、40.92%和52.33%;10~20 cm分别降低了15.97%、46.43%和58.33%。

图 4 不同季节氮沉降处理对土壤多酚氧化酶活性的影响Fig. 4 Effects of nitrogen deposition on soil polyphenol oxidase activity in different seasons
4 结论与讨论
土壤酶活性的季节变化可能是由土壤水分、温度、土壤养分及凋落物生态化学剂量比的直接作用和间接作用共同影响的。有研究结果表明,高温、降雨量丰富的季节土壤酶活性最高[13-14],而也有研究表明过高或过低的土壤温度和水分含量均会引起土壤酶活性的降低[15]。本研究结果显示,土壤脲酶和蔗糖酶活性最高值均出现在夏季,说明土壤水分和温度是这2种酶关键性的驱动因子。夏季为天山林区的雨季,林地内土壤湿润,凋落物分解加速[16-19],促进了微生物的繁殖和生长及酶活性的升高。李红琴[20]研究表明脲酶峰值出现在春季,这可能是由于春季微生物开始活动,但是此时植被生长刚刚开始,因此能量物质如新鲜的有机碳输入不足,从而导致对应的酶活性增强。本研究中,多酚氧化酶和过氧化物酶活性最高值出现在秋季,可能与秋季为天山云杉凋落物输入的高峰期有关。秋季大量新鲜凋落物中富含的可溶性有机质和纤维素等物质[17],可能刺激了氧化酶的活性,这对于促进凋落物高峰时产生的大量凋落物中的木质素降解和有机物质的腐殖质化具有重要作用[21],与罗飞[22]的研究结果基本一致。且氮沉降增加后脲酶和蔗糖酶活性季节变化特征未改变,而氧化酶活性季节变化规律发生了改变。
影响土壤酶活性对氮沉降响应差异的主要原因有土壤和植物初始氮水平、氮添加量、氮添加周期、外源氮类型[23]。目前研究普遍认为,长期施氮对氮“饱和型”森林土壤酶活性有显著的抑制作用,在氮增加的条件下,氮“缺乏型”不同森林生态系统土壤酶的反应有所不同[24-25]。宋学贵等[26]、涂利华等[8]及杜红霞等[27]的研究均认为氮沉降能够增加常绿阔叶林、苦竹(Pleioblastus amarus)林、连香树(Cercidiphyllum japonicum)林等林下土壤脲酶活性,与本研究结果一致。本研究中,尤其是夏季和秋季的表层土壤脲酶活性对氮沉降有正响应。这可能与土壤脲酶的主要来源及凋落物的分解有关。因为土壤脲酶主要来自植物根系分泌物[24],由于氮添加促进了植株的生长,进而导致根系分泌物的增加。而且夏季凋落物分解较快,根系分泌物也相应增加,导致土壤脲酶活性增加。另一方面,在氮缺乏的森林生态系统中,微生物与植物对氮的竞争激烈,微生物得不到充分的氮用于自身繁殖,当施氮量增加后,二者间的竞争逐渐减弱,微生物可利用的氮素增加,尤其是施高氮后,缓解了土壤微生物的氮素限制[25]。也有大量研究证明,土壤脲酶活性与土壤氮素水平为正相关。但也有研究表明,增氮抑制土壤脲酶活性,如周晓兵等[28]的研究发现,与对照相比,3.0、6.0 g/(m2·a)和24.0 g/(m2·a)处理使0~5 cm土层土壤脲酶活性降低了23.6%、37.1%和67.2%。这可能与施氮后,土壤无需矿化更多的有机物来满足植物和微生物生长的营养需求有关,也可能是施氮后导致土壤酸化,土壤脲酶活性降低。
多酚氧化酶和过氧化物酶是主要的木质素分解酶,施氮能够降低其活性[29]。如Saiya等[30]通过2 a的试验也发现,与对照相比,氮增加的处理多酚氧化酶活性降低了40%。这种增氮处理对2种氧化酶的抑制作用同样见于其他生态系统的研究[27]。目前,大量研究表明,多酚氧化酶和过氧化物酶是由白腐真菌产生,氮沉降增加能够抑制白腐真菌的活性,减少了这2种氧化酶的产量[31-32]。同时,已有分子层面的研究发现,这种负响应主要是由于施氮使得土壤中无机氮富集,氧化酶的基因表达受到抑制所造成的[13]。本研究结果表明,氧化酶的抑制作用在0~10 cm土层很显著,10~20 cm土层在低氮和中氮处理时不显著,仅在高氮处理抑制作用才显著。也有研究结果发现,氮添加对森林土壤氧化酶活性表现出无影响。如Ma等[33]在河北塞罕坝2种不同林龄华北落叶松进行2 a的氮添加试验,结果发现,氮添加对土壤多酚氧化酶活性没有显著影响。造成以上不同的结论的主要原因可能是生态系统中本底氮含量与氮输入量不同和土壤微生物群落结构不同。
土壤蔗糖酶可能影响土壤有机物质及营养元素的循环代谢、土壤-植物复合系统的碳、氮源汇动态及土壤质量[33]。涂利华等[8]研究发现,氮沉降促进了苦竹林土壤蔗糖酶活性。刘星等[34]的研究发现,氮沉降抑制了人工林中的蔗糖酶活性,对天然林没有影响,这可能是因为天然林的土壤本底氮含量略高于人工林的土壤本底氮含量。在本研究中,中氮和低氮处理对天山云杉林土壤蔗糖酶活性有促进作用,而高氮处理对其具有抑制作用。Eurasian Soil Science, 2016, 49(10): 1149-1160.
