广西云耳物种学名探究
2019-06-24祁亮亮韦仕岩蓝桃菊
祁亮亮 韦仕岩 蓝桃菊 郎 宁
(广西壮族自治区农业科学院微生物研究所,广西南宁530007)
云耳是云南、贵州和广西等地特色的木耳属食用菌,隶属于木耳目(Auriculariales),木耳科(Auriculariaceae)。木耳属真菌分布广泛,主要生长在阔叶树倒木或腐木上,偶见于腐朽的竹子上[1],是重要的食药用菌[2-4]。我国对木耳属的研究始于1939年,邓叔群记载了中国木耳属真菌6种[5];20世纪80年代后,国内学者相继发现报道了4个木耳属新种[6-8];近些年,基于形态学与分子系统学,诸多木耳属新种被发现[9-13]。截至目前,木耳属有效名称75个[13],中国有15个。云耳在广西多个市县有广泛栽培。
从宋代起,广西百色地区已开始人工栽培云耳。因此,云耳是广西特有的种质资源[14],一般发生在桂西、桂西北[15]。据记载,野生云耳大多发生于栎木腐木与麻栎林交界的混交林中,偶有在米椎(Castanopsisspp.)等其他树种上[14],多被描述为广西特有的木耳品种[16-19],学名常写作为Auricularia auricu-lar[14,19]或者A.auricular-judae[17],但缺乏有力证据。在宏观形态学上与云耳相似的物种多为近些年发表的新种或新记录种,如阔叶树木耳A.angiospermarumY.C.Dai,F.Wu﹠D.W.Li、黑木耳A.heimuerF.Wu,B.K.Cui﹠Y.C.Dai、小木耳A.minutissimaY.C.Dai,F.Wu﹠Malysheva、脆木耳A.fibrilliferaKobayasi等[12]。因此,笔者旨在通过比较云耳与其他相似种的形态学及分子系统学,完善广西云耳的形态学特征,明确广西云耳准确的物种名称及分类地位,对云耳的栽培育种及地方食用菌品种的保护具有重要意义。
1 材料与方法
1.1 供试材料
研究材料为笔者从广西天峨县(云耳712:GXMRI_YE1、GXMRI_YE2、GXMRI_YE3、)、南丹县(黑木 耳 916:GXMRI_ME1、GXMRI_ME2、GXMRI_ME3)、龙州县(GXMRI_0613、GXMRI_0627)采集的8个木耳属标本,标本存放于广西壮族自治区农业科学院微生物研究所(GXMRI)。
1.2 试验方法
1.2.1 形态学研究方法
采用传统形态学研究方法对新鲜及干标本进行宏观形态及微观显微结构的观察并记录。5%KOH溶液作为干标本的浮载剂,刚果红作为染色剂,Melzer试剂检验标本是否有淀粉质反应;测量30个成熟孢子的大小,得出所有孢子的长宽平均值,计算担孢子的Q值。颜色描述的依据是《Color standards and color nomenclature》[20]以及《Methuen handbook of color》[21]。
1.2.2 分子系统学研究方法
1.2.2.1 DNA提取、PCR扩增、测序
取干净无污染的木耳属干标本提取总DNA,提取方法依据DNA提取试剂盒SK8224(上海生工生物工程股份有限公司)使用说明。ITS序列的引物为通用引物[22],扩增反应体系为25 μL,包括1 μL模板,0.5 μL 正 反向引物 ,0.5 μL dNTP(10 mM),2.5 μL Tap Buffer,0.2 μL Tap 酶 ,20 μL ddH2O。PCR扩增反应条件为:95℃预变性3 min,94℃变性60 s,58℃退火30 s,72℃延伸1 min,共35个循环,72℃修复延伸10 min,4℃终止反应[23]。PCR产物送至上海生工生物工程股份有限公司测序。测序结果经验证后,上传至GenBank数据库。
1.2.2.2 数据处理及系统发育树的构建
基于NCBI数据库,将目标序列进行BLAST比对,下载其Ident值高于97%的序列组成矩阵。运用Bio Edit v.7.0.9[24]、CLUSTALX[25]和 PAUP 4.0[26]对 矩阵进行比对和编辑、Modeltest v.3.7[27]计算最适的碱基替换模型,MrBayes v.3.1.2[28]构建系统发育树。最终当运行100万代或分裂频率的平均标准偏差(Average standard deviation of split frequencies)低于0.01时停止运行,构建系统发育树。
2 结果与分析
2.1 云耳形态学
宏观形态(图1):子实体新鲜时胶质,不透明,耳状,偶不规则,具3~5 mm短小菌柄,边缘常具浅裂或偶具深裂,最宽处直径可达5.5 cm,厚0.50~1.46 mm,干后易碎,耳基及周围半透明(对光),耳缘不透明;不孕面浅黄褐色、深黄褐色,或木棕色(ⅩⅩⅤⅢ11″k),幼时具明显浅黄色柔毛,老后具黄棕色柔毛,干后浅灰棕色;耳缘不具柔毛,颜色不同,烟褐色、深红褐色,干后灰褐色;子实层光滑,具皱褶,幼时几乎与耳缘同色,基部色淡;老后子实层浅橙黄色(4A2),干后中部木棕色至淡黄棕色,边缘深灰色。
显微结构:担孢子腊肠形,无色,薄壁,光滑,具1或2个液泡(图2),非淀粉质,无嗜蓝反应,(3.0~5.5)μm×(7.0~12.0)μm,平均长度 L=10.18 μm,平均宽度W=4.20 μm,长宽比Q=2.42。担子棒状,3横隔,担孢子梗少见,(25.0~57.0)μm×(2.0~3.5)μm。子实层末端无拟囊状体。横切面具髓层,偶不明显,处于切面中间位置,无结晶体。柔毛单生或簇生,浅黄色,基部稍膨大,厚壁,具窄内腔,顶部钝圆,偶渐尖,大小为(46.0~99.0)μm ×(3.0~5.0)μm。菌丝具锁状联合,在5%KOH试剂中菌丝宽度为1.5~3.5 μm。

图1 云耳宏观形态

图2 云耳微观结构
在外观形态学上,云耳与阔叶树木耳、小孢木耳以及小木耳有很多相似之处,但是仍具有细微的差别(表1)。野生条件下,脆木耳很容易被误认为是云耳的野生物种,但是脆木耳新鲜时耳片较薄,易碎,呈半透明状。
2.2 云耳分子系统学
共得到26条ITS序列(表2),18条序列下载于GenBank,其余8条序列是自测序列。经对26条序列进行编辑整理后,矩阵中每条ITS序列碱基数为508个(包含空位)。最适碱基替代模型为TIMef,经过100万代的运行最终的分裂频率约为0.009,停止运行。贝叶斯方法构建的系统发育树如图3所示。

表1 云耳与相似种的形态学比较

表2 供试标本信息以及其GenBank登录号与采集地
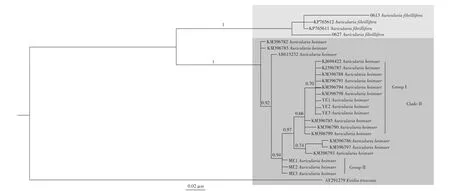
图3 基于ITS运用贝叶斯方法构建的系统发育树
运用贝叶斯方法,对26条ITS序列进行系统发育树的构建,其中15条序列是公认的黑木耳Auricularia heimuer,2条序列为脆木耳A.fibrillifera,1条序列为外群截形黑耳Exidia truncataFr.。由系统发育树可以明显看出,基于ITS构建的系统发育树将木耳属序列分为两个大的分支CladeⅠ和CladeⅡ。其中,0613、0627分布在CladeⅠ中,为脆木耳A.fibrillifera,属广西新记录种。YE1、YE2、YE3、ME1、ME2、ME3聚在CladeⅡ分支上,均为黑木耳。但是采自广西天峨县的云耳和南丹县的黑木耳并没有具在同一个小的分支上,云耳与中国及俄罗斯的黑木耳关系较近(GroupⅠ),分支支持率为0.70,且分支长相似,系统进化关系较近;而采自南丹的黑木耳标本单独形成一个小的分支(GroupⅡ)。因此可以确定采自天峨县和南丹县的6个标本均属于黑木耳A.heimuer,而并非是A.auricular或者A.auricularjudae。
3 小结与讨论
广西“百色云耳”具有悠久的栽培历史,距今已经1000余年。利用经典形态学研究方法,发现云耳在形态学上具有较为独特的特征,宏观上,其耳缘颜色、柔毛等与黑木耳不同;微观上,云耳的孢子、担子和柔毛均略小于黑木耳。野外采集调查时,脆木耳很容易被误认为是云耳,因为其子实体颜色十分接近,但新鲜脆木耳质地较薄,采摘容易开裂;脆木耳在光线稍暗的情况下,子实体近白色。在分子系统学方面得到的结果与吴芳研究结果相似[12]。云耳与来自俄罗斯及中国的部分菌株均聚在一个大的组内(GropeⅠ),但与俄罗斯的黑木耳在形态学上具有明显的区别,后者新鲜时具有棕黄色至红褐色的绒毛,干后黄绿色;担子和孢子也较云耳略大[29]。另外,黑木耳常规栽培种黑山A(KM396782)以及在广西的栽培种916(ME1-3)、云耳712(YE1-3)被分在不同的3个分枝上,由此可以推断,基于ITS序列构建的系统发育树能较好地对现有部分黑木耳品种进行区分[23]。另外,值得一提的是,AB615232是源于日本的黑木耳标本,在系统树中也能够明显的区分开来,可以推测出黑木耳种内具有复杂的遗传多样性。
综上所述,虽然在形态学上云耳与黑木耳存在部分差异,形成了广西地方特有的木耳栽培品种,但是综合形态学及分子系统学的观点支持将广西的云耳学名定为黑木耳Auricularia heimuerF.Wu,B.K.Cui﹠Y.C.Dai。另外,由于最初记载的云耳历史久远且未有形态学原始描述,并未获得野生子实体,故在子实体形态方面有待于今后进一步补充研究。
