家蚕黑色蚕突变体pB中apontic-like基因的结构与表达分析
2019-06-20王平阳谭静霞李亚萍孟祥川佟美金赵巧玲
蒋 迁 王平阳 谭静霞 李亚萍 孟祥川 佟美金 赵巧玲,3
(1江苏科技大学生物技术学院,江苏镇江 212018; 2广西壮族自治区蚕业研究院,广西南宁 530007;3中国农业科学院蚕业研究所,江苏镇江 212018; 4广西壮族自治区蚕业技术推广总站,广西南宁 530007)
昆虫体表形形色色的斑纹在拟态、躲避伤害、吸引异性、调节体温、免疫反应、表皮硬化、觅食、抵御紫外线等[1-10]许多生理生化方面起重要作用,是昆虫重要的生物学特征之一,同时也是昆虫在自然选择中能够繁衍下来的因素之一[11]。为了生存,许多昆虫拥有高度多样化的体色模式,即使在同一物种中,也呈现出极其不同的颜色,如非洲白凤蝶(Papiliodardanus)在一个简单的控制体色的基因座上,就拥有十多个等位基因[12]。在已有报道中,同一个基因(基因座)能控制多种不同体色表型的现象是很少见的,相关研究报道也较少[13],但是这类基因有助于对昆虫色素沉积机制的研究。
家蚕(Bombyxmori)作为鳞翅目昆虫的模式生物[14],拥有许多色素型突变体,非常适合作为研究色素代谢与斑纹形成机制的材料[15-17]。家蚕普通斑主要由3种形状的斑纹构成,即眼状纹、半月纹和星状纹,它们是由同一个基因座p控制的[18],而且将其定位到了第2连锁群上遗传距离大约3.0 cM的区间内[19],在这个基因座上,至少有15个等位基因,如素斑(plain,p)、极淡普通斑(lightest normal,pl)、普通斑(normal pattern,+p)、暗色蚕(moricaud,pM)、黑缟蚕(striped,pS)、腹条黑缟(ventral striped,pG)、黑色蚕(black,pB)、淡胸黑缟(whitish striped,pSw)、赛白斑(sable,pSa)、第2赛白斑(sable-2,pSa-2)等[18,20-22]。YODA等[23]通过图位克隆和基因功能分析,发现apontic-like(apt-like)基因是控制p基因座相关性状的关键基因,然后利用异位表达、RNA干涉(RNAi)、基因编辑(TELEN)等技术证明apt-like基因能引起细胞自主色素沉积,p基因座不同的着色模式是由于apt-like基因的表达水平不一样诱导黑色素合成路径上的基因表达量差异所引起的,其中pS是由于apt-like基因上发生单核苷酸多态性突变,引起1个氨基酸的替换,从而出现pS突变表型。控制pB突变体的主效基因pB是p基因座上另1个等位基因,pB突变体全身黝黑,各体节沿亚背线有若干白色波状带纹,属于单基因显性遗传,且纯合致死,采用杂合体保种[24]。
通过研究p基因座各等位基因的结构变化与表达模式,将有助于进一步研究apt-like基因在色素合成与斑纹形成中所扮演的角色,为昆虫斑纹形成机制的研究做贡献。本试验以pB突变体为研究对象,考察apt-like基因在pB突变体中的结构和表达量是否发生变化,从而证明pB突变体是否和pS突变体一样也是由于apt-like基因突变引起的,为进一步研究pB突变体色素的沉积机制奠定基础,也为apt-like基因的功能研究提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 供试家蚕品种 黑色蚕突变体pB(p/pB)由中国农业科学院蚕业研究所保存和提供,素蚕品种p(p/p)是从pB中分离而得,2个家蚕品种均在标准条件下饲养,发育至5龄第3天时解剖头部、表皮、中肠、血淋巴、卵巢、精巢、马氏管、气管、丝腺和脂肪体各组织备用。
1.1.2 主要试剂 DEPC(生物试剂)、琼脂糖(生物试剂)、溴酚蓝(生物染色剂)、蔗糖(生物试剂)、LB培养基(即用型粉剂,微生物用)和胶回收试剂盒,均为生工生物工程(上海)股份有限公司产品;RNA抽提液(RNAiso Plus)、DNase I、反转录试剂盒(PrimeScriptTMRT Master Mix)、荧光定量试剂盒[TB GreenTMPremix Ex TaqTM(Tli RNaseH Plus)(2×)]、蛋白酶K、RNase、DNA聚合酶(PrimeSTAR GXL DNA Polymerase和Ex Taq DNA Polymerase)、DL2000 DNA marker、dNTPs、T4 DNA连接酶和DH5α感受态细胞等试剂,均为宝生物工程(大连)有限公司(大连TaKaRa公司)产品;0.5 mol/L的EDTA(>99%,pH 为8.0)、3 mol/L的醋酸钠(分析纯,pH 为5.2)、冰醋酸(分析纯)、无水乙醇(分析纯)、75%的乙醇、10%的十二烷基硫酸钠溶液(化学纯)和氯仿(分析纯),均为国药集团化学试剂有限公司产品;1 mol/L的三羟甲基氨基甲烷[Tris(>99%,pH 为8.0)]和苯酚(>99%),均为北京拜尔迪生物技术有限公司产品。
1.1.3 主要仪器 TOMY SX-500型灭菌锅、GNP-9080型恒温培养箱,均为上海精宏实验设备有限公司产品;FRESCO 17型冷冻离心机、NANODROP1000型微量分光光度计,均为赛默飞世尔科技公司产品;LightCycler©96实时荧光定量PCR仪,为上海罗氏制药有限公司产品;G: BOX F3凝胶成像仪(SYNGENE)、2720 Thermal Cycle PCR仪,均为爱普拜斯应用生物系统贸易(上海)有限公司产品。
1.2 试验方法
1.2.1 试剂配制 DNA抽提液:取5 mL 1 mol/L,pH 为8.0的Tris,加入100 mL 0.5 mol/L,pH 为8.0的EDTA和25 mL 10%的十二烷基硫酸钠溶液,加去离子水定容至500 mL,灭菌,4 ℃保存备用;50×TAE:称取12.1 g Tris,量取2.85 mL冰醋酸和5 mL 0.5 mol/L,pH 为8.0的EDTA,加去离子水定容至50 mL,4 ℃保存备用;6× Loading Buffer:称取0.25 g溴酚蓝和40.00 g蔗糖,加去离子水定容至100 mL。
1.2.2apt-like基因的编码序列克隆 (1)黑色蚕pB、素蚕p的表皮组织提取总RNA。将表皮组织用液氮预冷的研钵研磨至粉末状,加入RNAiso Plus继续研磨至裂解液透明,将匀浆移入1.5 mL离心管中,然后参照RNAiso Plus说明书获得总RNA。(2)cDNA合成。总RNA测定浓度后,加入DNase I消化总RNA中残留的基因组DNA,然后参照PrimeScriptTMRT Master Mix说明书合成cDNA,cDNA测定浓度以后稀释至100 ng/μL。(3)apt-like基因的编码序列克隆测序。以apt-like作为引物(表1),pB和p的cDNA作为模板进行RT-PCR,产物电泳并进行胶回收,回收产物连接T载体并转化DH5α感受态细胞,挑取菌斑摇菌并进行PCR鉴定以后,送生工(上海)生物工程股份有限公司测序,测序结果进行比对以获得apt-like基因在2种家蚕中的编码序列。
表1apt-like基因克隆、定量和内参基因定量引物

引物名序列(5′→3′)作用apt-likeF: ATGTCAACGCGCAGTGCCR: TCATTTAGCGCTGGCCTTCTCapt-like基因编码序列克隆apt-qPCRF: TATCGAACACGCCTTTGAACCTGR: CATTCGTGCTTTATGTGATCGTCCAapt-like基因实时定量PCR分析apt-ge-nomeF: GGACGATCACATAAAGCACGAAR: TGAGTTTTGCAGCCATTACGAAapt-like基因第7外显子在基因组水平的克隆GAPDHF: TTCATGCCACAACTGCTACAR: AGTCAGCTTGCCATTAAGAG实时定量PCR用内参基因RPL3F: GAAGATGATCCGCTACTGTR: TATCCTTTGCCCTTGGTG实时定量PCR用内参基因
GAPDH为甘油醛-3-磷酸脱氢酶;RPL3为核糖体蛋白L3。
1.2.3apt-like基因第7外显子在基因组水平的克隆 (1)丝腺组织基因组DNA提取。将丝腺组织充分研磨后,加入0.6 mL的DNA 抽提液,冰上研磨,转入1.5 mL离心管,加入0.6 mL苯酚,充分混匀并离心,上清加入等体积的氯仿,充分混匀离心,上清加1/10体积的3 mol/L 醋酸钠(pH为5.2)和2倍体积的无水乙醇,充分混匀并离心,用1 mL 75%的乙醇洗涤沉淀,室温下风干,加适量去离子水溶解,测定浓度后稀释至100 ng/μL。(2)apt-like基因在基因组水平的克隆测序。参照“1.2.2apt-like基因的编码序列克隆”(3)中的方法,以apt-genome为引物(表1),基因组DNA作为模板对apt-like基因在基因组水平进行克隆测序,测序结果进行比对以获得apt-like基因在2种家蚕中的最后1个外显子序列。
1.2.4apt-like基因的组织表达分析 (1)黑色蚕pB、素蚕p的5龄第3天各组织总RNA提取。 参照“1.2.2apt-like基因的编码序列克隆”(1)中的方法获得黑色蚕pB和素蚕p的5龄第3天各组织总RNA。(2)cDNA合成。参照“1.2.2apt-like基因的编码序列克隆”(2)中的方法合成cDNA,cDNA测定浓度以后稀释至100 ng/μL。(3)apt-like基因的实时定量分析。以apt-qPCR作为定量引物(表1),pB和p各组织的cDNA作为模板进行qRT-PCR,以家蚕GAPDH和RPL3为内参基因(引物见表1),实时定量PCR反应体系(20 μL)为cDNA模板1 μL,上下游引物(表1)各0.5 μL(10 μmol/L),TB GreenTMPremix Ex TaqTM(Tli RNaseH Plus)(2×)10 μL,去离子水 8 μL。反应程序为95 ℃预变性10 min,随后40个3步循环(95 ℃变性10 s,57 ℃退火10 s,72 ℃延伸10 s),所得数据采用2-ΔΔCT法[25]计算apt-like基因的相对表达量。
2 结果与分析
2.1 apt-like基因的编码序列克隆
在pB和p中均获得大约950 bp左右大小的条带(图1),未出现显著差异,进一步对PCR产物进行克隆测序,结果表明apt-like基因编码序列在pB和p中序列完全一致,并未出现结构上的变异,与数据库中的apt-like基因序列相比有4个碱基的替换,但是其翻译的氨基酸序列完全一致,表明apt-like基因编码序列在黑色蚕突变体pB中并未出现结构上的变异。
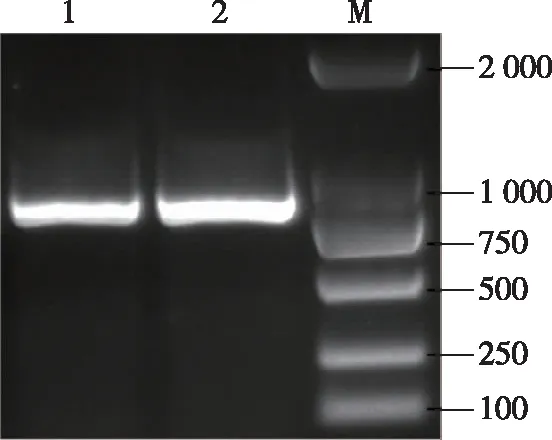
1. apt-like基因编码序列在素蚕p中的PCR产物,2. apt-like基因编码序列在黑色蚕pB中的PCR产物,M.DL2000 DNA marker。图1 家蚕apt-like基因编码序列的克隆
2.2 apt-like基因在基因组水平的克隆
编码序列克隆结果表明,apt-like基因在黑色蚕突变体pB和素蚕p中没有出现结构上的差异,且在黑缟蚕突变体pS中是apt-like基因的最后一个外显子出现结构变异,因此对黑色蚕突变体pB和素蚕p中apt-like基因的最后一个外显子进行克隆(图2),结果未出现显著差异,进一步测序结果表明apt-like基因的最后一个外显子在pB和p中序列完全一致,进一步证明黑色蚕突变体pB中apt-like基因未发生结构变异,且由于黑色蚕突变体pB和黑缟蚕突变体pS突变位点不一样,因此apt-like基因并未在最后一个外显子出现单核苷酸多态性突变。

1. 基因组上apt-like基因最后一个外显子在素蚕p中的PCR产物,2. 基因组上apt-like基因最后一个外显子在黑色蚕突变体pB中的PCR产物,M. DL2000 DNA marker。图2 家蚕apt-like的基因组鉴定PCR产物
2.3 apt-like基因的组织表达分析
经过在mRNA和基因组水平对apt-like基因编码序列进行克隆测序,发现编码序列在结构上并未发生变化,于是利用实时定量PCR对apt-like基因的表达量进行分析。分别用黑色蚕突变体pB和素蚕p的5龄第3天的表皮、头部等10个组织器官进行定量分析(图3)。结果显示,apt-like基因在表皮、头部和中肠中表达量较高,除在血淋巴中表达量没有显著差异外,在其他9个组织中,apt-like基因的相对表达量均是素蚕p显著或极显著高于黑色蚕突变体pB(图3)。除血淋巴外,黑色蚕突变体pB的其他组织apt-like基因与素蚕p相比都是显著下调表达,其中在中肠、卵巢、气管和脂肪体中下调倍数超过2倍。这些结果表明,在黑色蚕突变体pB中apt-like基因的表达量受到了显著影响,可能在黑色蚕突变体pB突变性状的形成中起作用。

*表示差异显著(P<0.05),**表示差异极显著(P<0.01)。图3 家蚕apt-like基因在黑色蚕突变体pB和素蚕p的5龄第3天各组织的相对表达量
3 讨论
显性遗传是指同源染色体其中一个突变即可引起突变表型的遗传模式,显性纯合或者杂合都表现为突变性状,只有隐性纯合时才表现出野生型性状。在家蚕中也有许多显性遗传的突变体,楔形眼纹(Wes)是一种显性遗传,纯合(Wes/Wes)致死,杂合(+/Wes)可以生存,表现出突变表型,该突变基因位于家蚕第6连锁群上[26]。与Wes类似,短体蚕(Sq)[27]、竹蚕(Bo)[28]、裸蛹(Nd)[29]等都是显性遗传突变体。如果显性突变纯合致死(胚胎致死),就要用杂合体作为突变体进行研究,而杂合体有一半是正常野生型基因,为突变基因克隆带来麻烦,当突变基因因突变无法克隆时,杂合体往往都是克隆出了正常的基因,因此最好的办法是用显性纯合的胚胎进行克隆[30]。
黑色蚕突变体pB由于胚胎致死,使用未孵化蚕卵(大部分应为纯合致死)无法获得足量可用的总RNA,可能正常蚕孵化后纯合致死卵已经死亡且内容物降解,为此,在用杂合体进行克隆时,加大了检测样本量(n>40),在pB和p中未发现apt-like基因编码序列有结构差异,然而表达量检测发现apt-like基因在pB和p之间有差异,且几乎所有检测的组织(血淋巴除外)中apt-like基因在pB中的表达量相对于在p中的表达量显著下调,而且在突变体pB(p/pB)中还有一半正常基因,暗示突变基因pB几乎不表达,说明apt-like基因在黑色蚕突变体突变性状的形成中是起作用的,而且有可能起了关键作用。
经克隆测序,未发现apt-like基因编码序列在pB(p/pB)中发生结构变异,可能是由于发生突变的染色体出现了大片段的缺失、大片段的插入或者表达量极低等情况而导致PCR获得的只是正常染色体的表达产物,也可能是由于发生突变的染色体上apt-like基因的编码序列没有发生结构变异,而是非翻译区或者启动子发生了结构变异。后续我们一方面将对apt-like基因的非翻译区和启动子进行研究,另一方面还将尝试使用显性纯合畸形胚胎克隆apt-like基因。
