粤北阔叶人工林和次生林植物多样性与土壤理化性质相关性研究
2019-06-18闫玮明孙冰裴男才王旭李非凡罗鑫华邹滨
闫玮明,孙冰*,裴男才,王旭,李非凡,罗鑫华,邹滨
1. 中国林业科学研究院热带林业研究所,广东 广州 510520;2. 广东乐昌杨东山十二度水省级自然保护区管理处,广东 乐昌 512231;3. 乐昌大瑶山省级自然保护区管理处,广东 乐昌 512231
中国人工林发展迅速,已成为世界人工林保存面积最大的国家(陈幸良等,2014),缓解了中国木材利用紧张的问题。近年来,人工林集约化经营模式对生态系统干扰增强,带来经济效益的同时,出现林分质量不高、生产力水平低、稳定性差等问题,其生态效益越来越受到重视(刘世荣等,2017)。模拟地带性森林群落物种组成,对人工林进行适宜抚育间伐措施,是全面推进人工林可持续经营的重要措施之一,因此研究人工林与天然林的群落组成差异尤为重要(陈幸良等,2014)。人工林植物多样性主要通过林下植被反映,林下植被是人工林发展生态效益的重要部分(Mori et al.,2004),适当丰富能促进人工林和天然次生林乔木层生长繁殖、提高目标树种产量(Thomas et al.,1999;卢志军等,2013),有利于森林结构塑造,维持生态系统可持续发展(李敏敏等,2018)。林下植被与土壤状况密切相关。土壤是群落发生和发展的基础,较好的土壤能够协调水分、空气和营养物质的关系,直接或者间接影响林木的健康发展,是生态系统中多种生态过程必不可少的载体(Castello et al.,2011;Wingfield et al.,2015)。
目前,林下植被的研究从单一树种人工林扩展到不同植被类型人工林与天然次生林林下植被差异分析。天然次生林与人工林林下物种多样性对比研究因林分类型、地理环境差异而呈现3种结果:其一,天然林群落层次比人工林复杂,物种多样性高,如珠海市红树林天然群落物种多样性优于人工林(张晓君等,2014);其二,人工林与天然林群落结构组成无明显区别,经过一定时间恢复的木荷人工林,其物种组成和群落结构均接近天然林(胡喜生等,2007);其三,人工林灌木层多样性低于天然林,而草本层丰富度高于天然林(朱锦懋等,1996)。究其原因,林下土壤理化性质差异发挥了重要作用。原始林、次生林、人工林的土壤有机碳、总氮、速效氮含量依次递减(魏彦昌等,2007);与人工林相比,天然林土壤结构状况更佳,保水抗蚀性较强(何毓蓉等,2005);天然林土壤密度低,全氮、全磷、水解氮、速效钾均高于其他林分(王新宇等,2008)。
粤北山地是广东省最大的森林资源库和重要生态屏障。近年来,很多研究关注粤北次生林生物多样性维持机制,特别是 2008年冰雪灾害后常绿阔叶林的恢复和更新(叶万辉等,2008)。森林群落多样性与土壤理化性质紧密相关,二者之间是否正相关性未有定论(崔宁洁等,2014;姚俊宇等,2018)。因此,选择粤北地区红锥(Castanopsis hystrix)乐昌含笑(Michelia chapensis)和深山含笑(Michelia maudiae)(二者同属木兰科,故下文中将其视为一种林分,称含笑人工林)3种人工林及天然次生林为研究对象,分析人工林与天然次生林林下植被物种组成差异,了解林下植物多样性与土壤理化性质的关联效应,分析林下植物多样性与土壤理化性质的相关性等科学问题,以期指导粤北人工林营造技术,为更好发挥人工林生态效益与经济效益提供科学依据。
1 研究区域概况
试验样地位于广东省乐昌市龙山林场(131°11′58″E,45°35′53″N)。龙山林场地处乐昌市廊田镇,属于中亚热带至南亚热带的过渡地段,中亚热带季风气候,常年温暖,雨量充沛,湿度较大,年平均气温19.6 ℃,年平均日照时数 1499.7 h,无霜期300 d。干湿季明显,4-9月为雨季,10月至翌年3月为旱季,年均降雨量1522 mm,雨热同期,光、热、水资源丰富,土壤为花岗岩发育的红壤,独特的地形和适宜的气候条件使得当地物种多样性较高。植被主要类型为中亚热带常绿阔叶林,其次为常绿落叶阔叶林以及针阔叶混交林、竹林、山顶矮林、高山草甸和各类人工林等(刘旭平等,2018)。
2 研究方法
2.1 样地设置
基于对研究区龙山林场现有森林资源的详细勘察,选取土壤类型均为红壤的红锥(Castanopsis hystrix)人工林、乐昌含笑(Michelia chapensis)人工林、深山含笑(Michelia maudiae)人工林和天然次生林,每种林分分别设立3个标准样地,样地大小为30 m×40 m,共设置9个样地,样地基本情况见表 1。其中,红锥人工林与含笑人工林造林时间均为2002年,初始密度2 m×2 m,种植5 a后,为培育大径材,进行抚育间伐,定植密度 800 plant·hm-2;天然次生林为1958年砍伐后自然恢复的亚热带常绿阔叶林,林龄60 a。
2.2 调查方法
于 2018年 4-6月对样地进行调查取样,用GPS记录每个样地中点经纬度、海拔等。采用相邻格子法将每块样地划分为12个10 m×10 m样方,调查内容包括:每个样方中胸径(DBH)≥5 cm乔木的种名、胸径、树高、冠幅、枝下高。每个样地选取5个10 m×10 m样方,调查所有灌木(包括胸径DBH≤5 cm的乔木)种名、胸径、株丛数、高度、盖度;在每个灌木样方对角线上各设置3个2 m×2 m的小样方,调查草本植物及乔木、灌木的幼苗的种名、胸径、株丛数、高度、盖度(王丽娟等,2017;张珂,2017;许涵等,2009)。
2.3 土壤理化性质测定
2.3.1 土壤样品采集
采用五点取样法,选取样地的相应位置(即样方对角线交点,再选择对角线上与交点距离相等的四个点,共五点)挖土壤剖面;用100 cm3的环刀分别采集0-20、20-40 cm深度的土样,3次重复,每个样地取 15个环刀用于土壤物理性质的测定;另外逐层取散装土用于土壤化学性质测定,每三点土样混匀后用四分法取对角线土样作为1份测定土样,3次重复(张珂,2017)。
2.3.2 土壤理化性质的测定
土壤密度、土壤质量含水量采用环刀(100 cm3)取样分析法测定(国家林业局,1999)。
土壤化学性质:土壤pH值采用电位法测定;土壤有机质采用重铬酸钾氧化外加热法测定;全氮采用凯氏定氮法测定;全磷采用 NaOH碱溶-钼锑抗比色法测定;速效磷采用0.5 mo·L-1NaHCO3浸提-钼锑抗比色法测定;全钾、速效钾采用火焰光度计法测定;阳离子交换量采用EDTA-铵盐快速法测定;铵态氮采用2 mol·L-1KCL浸提-靛酚蓝比色法测定;硝态氮采用双波长紫外分光光度法测定(鲍士旦,2000)。

表1 样地基本情况Table 1 Characteristics of the nine plots
2.4 分析方法
2.4.1 物种重要值计算
乔木层重要值:

林下灌木、草本层重要值(马淑琴等,2017):

式中,IV是物种重要值,(Important Value);RA为相对多度(Relative Abundance);RF为相对频度(Relative Frequency);RBA 为相对断面积(Relative Basal Area);RC为相对盖度(Relative Coverage)。
2.4.2 物种多样性计算
生物多样性的衡量指标采用 Shannon-Wiener指数,Simpson优势度指数,Pielou均匀度指数,Margalef丰富度指数。公式分别如下(马克平等,1994):
Shannon-Wiener指数:

Pielou均匀度指数:

Margalef丰富度指数:

式中,S为物种总数;Pi为物种 i植物个体数占群落中总个体数的比例;N为样地中所有个体数的总和。
2.5 数据处理
采用SPSS 19.0中的单因素方差分析不同林分物种多样性指数和土壤理化性质的显著性检验,Pearson相关性分析物种多样性与土壤理化性质的相关性。运用Excel 2016进行数据统计和制图。
3 结果与分析
3.1 不同林分物种组成差异
红锥林林下植被共计33科40属41种,灌木层植物 11科 14属 14种,包括杜茎山(Maesa japonica)、杉木(Cunninghamia lanceolata)和粗叶榕(Ficus hirta)等,草本层植物29科32属32种,优势种有红锥幼苗,石楠藤(Piper puberulum)等;含笑人工林林下植被共计34科40属43种,灌木层植物7科8属9种,粗叶榕占绝对优势,草本层植物 33科 33属 36种,优势种有阔片短肠蕨(Allantodia matthewii)、半边旗(Pteris semipinnata)、石楠藤、火炭母(Polygonum chinense)等;天然次生林中,林下植被共计35科48属61种,灌木层植物11科15属19种,主要包括黧蒴锥(Castanopsis fissa)、广东润楠(Machilus kwangtungensis)、鸭公树(Neolitsea chuii)等,草本层植物33科40属47种,优势种有华山姜(Alpinia chinensis)、石楠藤等。2种人工林林下优势种的相似性较高,天然次生林灌木层中出现了区别于人工林的植物种类,优势种植物以乔木层树种幼苗居多,人工林乔木层中占优势的杜茎山和粗叶榕消失;草本层优势种中蕨类植物少于人工林(表 2,表3),说明在天然次生林中,杜茎山、粗叶榕等在演替初期是优势种,随着演替的进行逐渐消失并最终被树种幼苗替代。
红锥人工林林下出现了数量可观的红锥幼苗,是草本层中的优势种;含笑人工林林下并未出现与乔木层对应的植物;天然次生林中,林下出现了与乔木层相对应的树种,如新叶木姜子(Neolitsea aurata)、鸭脚木(Schefflera heptaphylla)、野柿子(Diospyros kaki)、木荷(Schima superba)、广东润楠等。说明红锥人工林与天然次生林随着时间的发展能够实现自我更新,有利于群落更新,维持相对稳定的群落结构。
3.2 不同林分林下植被多样性差异
对3种林分灌木层、草本层的物种多样性指数进行比较,结果如图1、图2。
灌木层中,天然次生林Shannon-Wiener指数、Simpson优势度指数、Margalef丰富度指数3者呈现相同的变化趋势,均表现为天然次生林最高,其次为红锥人工林,木兰科人工林最低,与3种林分灌木层包含植物种数的大小关系一致;草本层的物种多样性指数Shannon-Wiener指数和Pielou均匀度指数呈现红锥人工林>木兰科人工林>天然次生林,Margalef丰富度指数呈现天然次生林>木兰科人工林>红锥人工林。
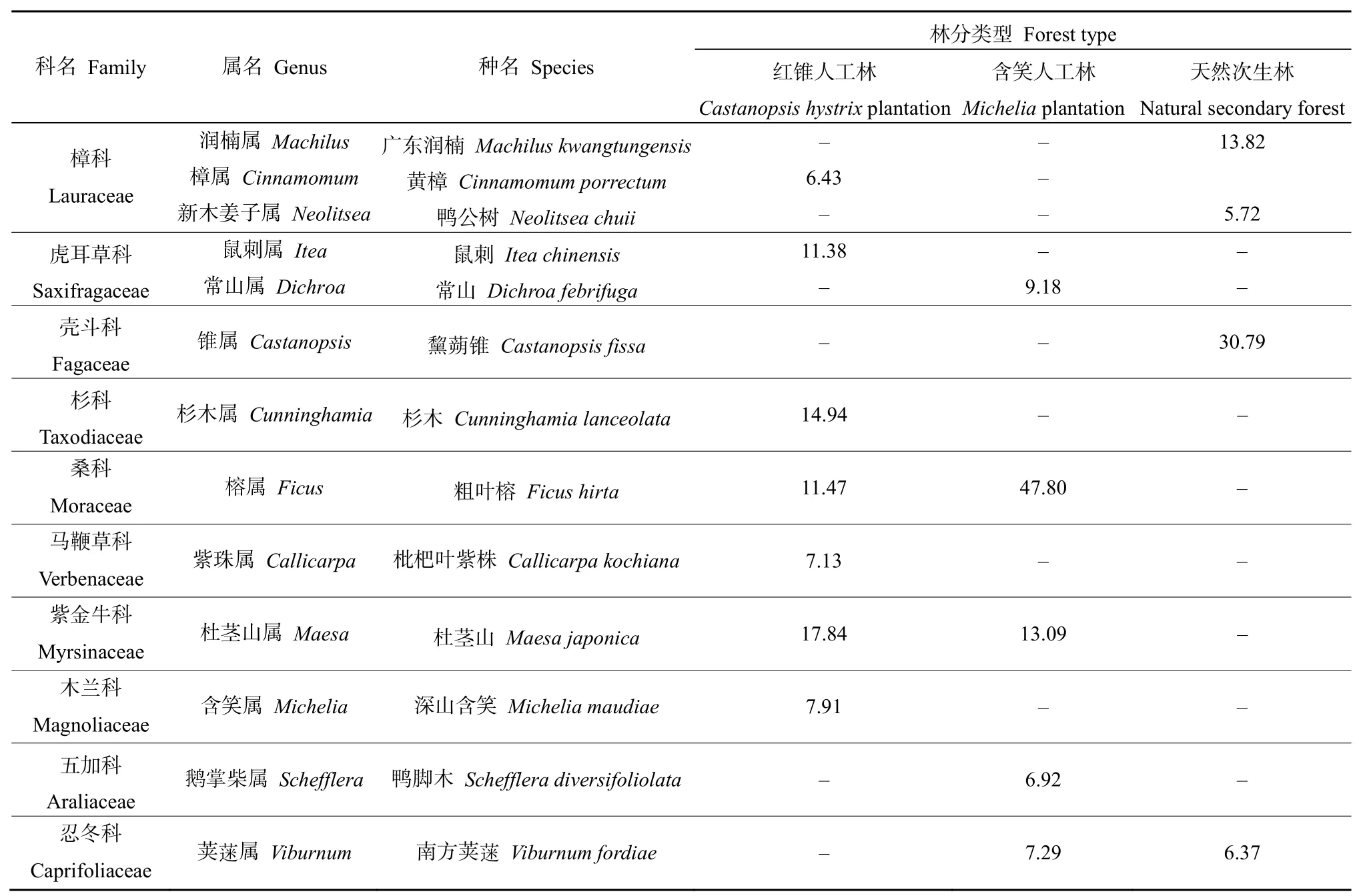
表2 灌木层主要物种组成及其重要值Table 2 Main species composition and importance values of the shrub layer

表3 草本层主要物种组成及其重要值Table 3 Main species composition and importance values of the herb layer

图1 不同林分灌木层物种多样性指数Fig. 1 Species diversity index of the shrub layer in different forests
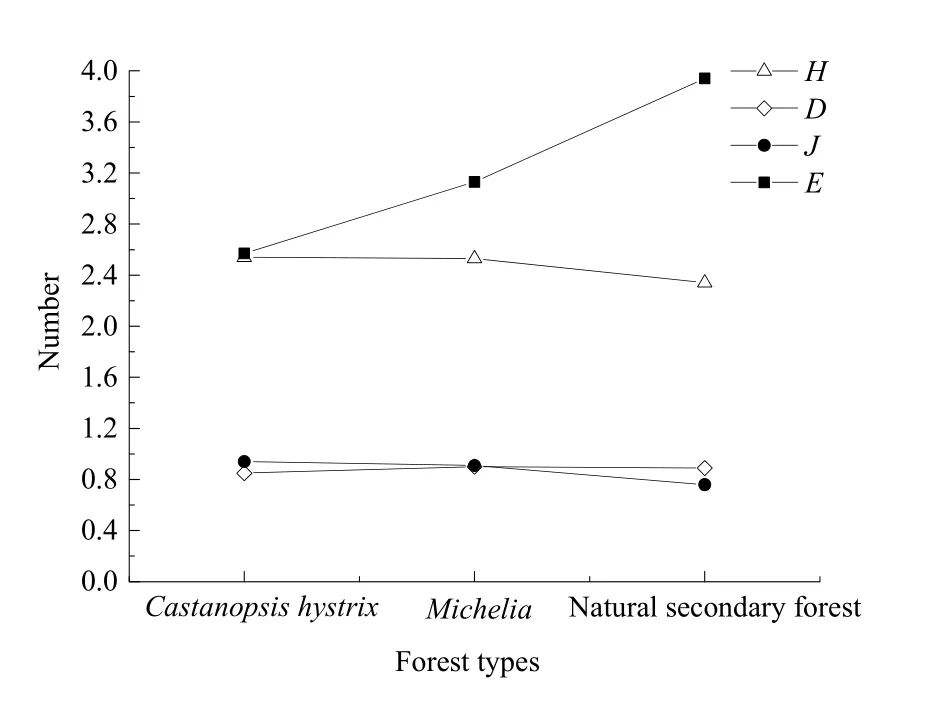
图2 不同林分草本层物种多样性指数Fig. 2 Species diversity index of the herb layer in different forests
3种林分灌木层与草本层物种多样性指数均表现为草本层>灌木层,均未满足自然生态系统各层次物种多样性指数灌木层>乔木层>草本层的格局,均处于发展中。
3.3 不同林分土壤理化性质
3种林分的土壤理化性质测定结果见表4。
土壤理化性质含量随土层深度的变化呈现规律性变化。土壤密度、全钾随土壤深度的增加而增加;质量含水量、有机质、全氮、全钾、速效磷、速效钾、阳离子交换量、铵态氮、硝态氮均随土层深度的增加而减小。
土壤理化性质因林分类型不同而存在显著差异。土壤物理性质方面:土壤密度与土壤质量含水量在不同林分间差异显著(n=45,P<0.05)。不同土层深度土壤密度表现为天然次生林>含笑人工林>红锥人工林,质量含水量表现为红锥人工林>含笑人工林>天然次生林,说明人为干扰可能导致人工林土壤密度减小,含水量增加。土壤化学性质方面,3种林分土壤pH值在4.17-4.38之间,呈酸性。土壤全氮、全磷、全钾、阳离子交换量、铵态氮在不同林分类型差异显著(n=45,P<0.05)。分析土壤各理化性质指标发现,速效钾和有机质含量的质量分数较大,且表现为红锥人工林>含笑人工林>天然林,表明二者含量易受人为干扰的影响。速效磷在3种林分的含量大小与其他土壤化学性质相反,表现为天然次生林>含笑人工林>红锥人工林,可能与其他指标存在负相关性,硝态氮各林分差异不显著(n=45,P>0.05)。

表4 龙山林场不同林分土壤理化性质Table 4 Soil physical properties in different forests in Longshan Forest Farm
3.4 生物多样性与土壤理化性质的相关性分析
植物群落物种多样性指数与土壤理化性质关系密切(表 5),灌木层中,0-20 cm 土层内Shannon-Wiener指数、Margalef丰富度指数与速效钾呈显著负相关(P<0.05),Pielou均匀度指数均与全钾呈极显著负相关(P<0.01);20-40 cm土层,Pielou均匀度指数均与全钾呈极显著负相关(P<0.01)。草本层中,Shannon-Wiener指数在不同土壤深度均与全钾呈显著负相关(P<0.05);Pielou均匀度指数在不同土层均与质量含水量、田间持水量、毛管孔隙度、有机质等呈显著正相关(P<0.05),与土壤密度、pH值、全钾、速效磷呈显著负相关(P<0.05)。
研究表明,3种林分中,灌木层物种多样性可能与土壤中钾含量有关,与土壤物理性质的相关性不显著,草本层物种多样性与土壤含水量、密度、全钾、速效磷、有机质均呈显著相关。草本层物种多样性指数与土壤理化性质相关性更强。
4 讨论
群落物种多样性是反映一定空间范围内物种丰富度和物种均匀度的重要指标,与群落中物种数量及其分布频率密切相关,能够体现群落结构类型、组织水平、发展阶段和稳定程度(程真等,2015;马志波等,2016)。本研究中,天然次生林比人工林林下植被物种数丰富,单科单属生活型植物比例降低,原因可能是人为干扰破坏了人工林林下物种组成,改变林分结构,从而影响生物多样性甚至群落演替进程(方升佐等,2012)。Nagaike et al.(2006)将人工林与天然更新林分进行对比,认为经过适度人为除草和间伐的人工林中,乔木树种和依靠鸟类散播种子的植物种类和丰富度降低。相关学者认为,人工林物种多样性低的原因之一是人工林本身的局限性,经营措施是造成物种多样性低下的主要原因(Brockerhoff et al.,2008;Aubin et al.,2008),此外,还受到来自邻近天然林的种子的影响(Onaindia et al.,2009)。为了平衡人工林的环境和经济效益,可通过混交造林(Hartley,2002;Paquette et al.,2010)、阔叶树种造林(Aubin et al.,2008)、延长轮伐期(Munro et al.,2009)、选择地带性植被(夏艳菊等,2018)等手段,丰富人工林物种多样性。
土壤是气候、母质、植被、地形长期综合作用下的产物,并随着群落演替不断发生变化(康冰等,2010)。土壤理化性质的差异决定了群落结构的不同,而不同的植物因生长活动、凋落物的分解等直接或间接影响土壤的理化性质(叶万辉,2000;贺金生等,2010;魏晨辉等,2015)。本研究中,天然次生林全钾、速效磷含量明显高于2种人工林,说明天然次生林土壤有较强的养分循环和富集功能,人工纯林经营强度大,群落组成单一,能够归还土壤的凋落物数量和养分量少,强烈的人为干扰,林下灌草被清除,土壤养分被不断消耗,是人工林养分含量低的主要原因(张贾宇等,2018)。但天然次生林全氮、全磷含量较低,原因是天然次生林地表地表凋落物的分解是土壤N的主要来源,天然次生林地表凋落物较丰富,一定程度上影响有机质分解和循环,故土壤中氮含量较低(苗娟等,2014);也可能是亚热带森林中,林龄较大的林分土壤预期溶解有机氮的损失高于林龄低的林分(fang et al.,2009)。康冰等(2010)研究发现“移针引阔”措施能够通过增加物种多样性来改变群落养分循环,形成良好土壤养分循环机制,丰富土壤养分含量。因此,提高人工林林下植被多样性,丰富人工林群落结构,经过缓慢的发展,其土壤养分会有明显改善,从而更有利于林分内部养分循环,增强群落稳定性。

表5 物种多样性多样性与土壤养分的相关系数Table 5 Correlation coefficients between diversity indices and soil nitrogen
相关性分析发现土壤理化性质与物种多样性之间存在对应关系,土壤氮、磷含量是土壤肥力的重要指标(Timothy et al.,2013),有效氮是限制植物生长的重要因子(Sigurdsson,2001),全磷是南亚热带森林的限制因子(刘兴诏等,2010),土壤 pH值和有效磷是群落中物种多样性的影响因子(Xu et al.,2016)。本研究结果表明,有机质含量、全氮、速效磷与林下多样性指数存在相关关系,可能是影响其林下植被生长的关键因素;草本层相对于灌木层而言,与土壤理化性质的相关性更强。出现该现象可能是因为草本层抗干扰能力弱,对土壤因子较为敏感,由此可以推断,土壤理化性质对于草本植被的影响更大。目前有关土壤理化性质与林下植被物种多样性指数的相关性未有统一定论,有待进一步研究(崔宁洁等,2014;姚俊宇等,2018)。
5 结论
相较于同一地带的天然次生林,粤北地区16 a生人工林林下物种成分简单,物种多样性较低,乔木层优势种的幼苗稀少,导致人工林自然更新能力低下,严重制约了森林生态服务能力和林分生产力,需要加强补植、套种等人工干预措施,调整人工林物种组成和林分结构。
氮、磷、钾含量和土壤pH是制约林下植被物种多样性的关键因子,特别是土壤中钾含量与乔木层幼苗的萌发和生长关系密切,对于生物多样性维持及次生林和人工林的顺行演替有深远影响,需在今后的森林立地管理和土壤保育中予以特别关注。
