胚挽救技术在我国无核葡萄育种中的应用及展望
2019-06-01赵艳卓陈展牛早柱魏建国牛帅科杨丽丽
赵艳卓,陈展,牛早柱,魏建国,牛帅科,杨丽丽
(河北省农林科学院石家庄果树研究所,河北 石家庄 050061)
葡萄(Vitis vinifera)是世界四大水果之一,栽培遍及五大洲,鲜食葡萄因粒大味美而深受消费者的喜爱。无核葡萄外形美观,品质优良,食用方便,在鲜食市场和制干领域均占据重要地位。目前,无核葡萄品种是我国各葡萄产区的主推品种[1],其中,大部分为国外育成品种,国内育成品种较少。市售鲜食无核葡萄中大多是将有核品种经过无核化处理后得到的,如玫瑰香、巨峰、藤稔、红地球、醉金香、阳光玫瑰等,天然无核品种只占一小部分。随着国际果品安全进程的不断推进,利用化学物质诱导葡萄无核化势必要被应用绿色安全的无核品种所取代。因此,加大无核葡萄育种力度,选育大粒、优质且具香味的鲜食无核葡萄品种成为当今葡萄育种的主要目标[2,3]。
无核葡萄育种途径有芽变选种、杂交育种、多倍体育种、胚挽救育种、分子标记辅助育种等,其中杂交育种是无核葡萄育种的主要手段。传统的杂交方式只能以有核葡萄为母本,依靠传递力强的无核父本来传递无核性状,利用这种方式选育无核葡萄品种效率很低,杂种后代中无核的几率一般不超过15%[4],且选育过程时间长。但这一状况在1982年因Ramming等创立无核葡萄胚挽救(embryo rescue)技术而改变。胚挽救技术是运用植物组织培养技术,对假单性结实或其他发育有障碍的受精胚或胚珠,在其合子胚败育之前进行离体培养,阻止幼胚败育,利用胚乳及培养基中的营养使其发育成充实的胚,最终形成完整的植株[5]。该技术使无核葡萄×无核葡萄的杂交方式成为现实,是目前无核葡萄育种研究中发展最快的新兴实用技术,推动了无核葡萄的育种进程。我国自20 世纪90年代开始无核葡萄胚挽救技术研究,现已在无核葡萄种质创新[4,6~15]、三倍体无核品种选育[8,10,11,16~20]、抗性品种选育[21~26]等方面广泛应用,同时对影响胚挽救的不同因素[3,9,21,24,27~34]进行了研究。
1 胚挽救技术在无核葡萄育种中的应用
1.1 杂交育种
杂交育种是葡萄育种中使用最广泛的技术。我国培育的葡萄品种中,约有70%是通过杂交获得的[5,35]。与传统育种相比,胚挽救技术因获得的子代无核率高,且育种周期短,而被广泛应用于葡萄无核种质的创制。通过葡萄种内杂交,或者与葡萄属其他种间杂交,如与圆叶葡萄(V.rotundifolia)、山葡萄(V.amureusis Rupr.)、刺葡萄(V.davidii Foex.)、蘡薁葡萄(V.thunbergii Sieb.)、欧山杂种葡萄等杂交,我国葡萄科技工作者获得了许多具有不同特性的无核葡萄新种质[7~9,24,26,36]。如,为培育无核、香味葡萄新品种,多位学者以无核葡萄品种做母本,二倍体或四倍体玫瑰香型葡萄或优系做父本,配置不同杂交组合,获得了一批胚挽救株系[7,9,29,37,38]。
1.2 三倍体育种
目前获得三倍体葡萄的有效途径是二倍体与四倍体进行有性杂交,但由于染色体的不均衡性,导致胚在发育早期即发生败育,或获得的杂交种子生活力低,造成育种效率低下。利用胚挽救技术可阻止三倍体杂交幼胚的早期败育,获得三倍体植株,从而提高育种效率。李世诚等[39]较早(1998年)应用胚挽救技术得到了三倍体葡萄植株,随后,众多学者也通过胚挽救技术得到了三倍体葡萄,并对三倍体育种中胚挽救技术的影响因素以及三倍体植株的鉴定进行了研究[17,20,39~43]。近期,田淑芬等[29,37,38]以二倍体无核品种希姆劳特、红宝石无核、奥迪亚无核为母本,与四倍体品种巨玫瑰杂交,并对影响胚发育及成苗的因素进行了研究,经倍性检测获得‘希姆劳特×巨玫瑰’的三倍体幼苗1 株。
在三倍体育种方面,多数研究是在欧亚种葡萄与欧美杂种葡萄间进行,仅有少数研究是在欧亚种与欧山杂种[19]、刺葡萄[44]间进行。唐冬梅[8]对不同倍性杂交组合后代进行分析后认为,2x×4x 杂交方式较4x×2x 杂交方式能得到更多的杂交后代,但4x×2x 杂交方式较2x×4x 杂交方式获得的杂交后代中三倍体的比例高。
1.3 远缘杂交育种及抗性育种
目前生产上广泛栽培的无核葡萄品种多数属于欧亚种,其果实品质优良,但抗病性和抗逆性较差。葡萄属其他种则具有不同的品质特性以及优秀的抗病、抗逆性,如圆叶葡萄对根瘤蚜完全免疫,我国野生葡萄中的山葡萄具有极佳的抗寒性,但由于亲缘关系较远,造成传统的育种方法效率低。而利用胚挽救技术可以提高培育抗病、抗逆无核葡萄新品种的效率。
在远缘杂交方面,1988年Goldy 等首次利用胚挽救技术将欧亚种无核葡萄与圆叶葡萄远缘杂交,探究了获得葡萄新种质的可能性。在国内,潘学军等[43]首次报道了利用圆叶葡萄与欧亚种葡萄杂交,成功获得亚属间杂种苗19 株。随后,其他学者陆续报道了蘡薁葡萄泰山-2[23,26],刺葡萄塘尾[23,25]、紫秋[44]、雪峰[19],秋葡萄江西-2[23],燕山葡萄[45],秦岭葡萄平利-5[45]等与欧亚种或欧美杂种葡萄进行杂交的研究。
在抗逆性方面,除少数学者采用蘡薁葡萄泰山-2、刺葡萄塘尾、秋葡萄江西-2 等,多数学者采用抗寒性极强的山葡萄如黑龙江实生、左山74-1-326、双优等或抗寒性较强的欧山杂种北醇、北冰红等与欧亚种葡萄杂交,以提高欧亚种葡萄的抗寒性[8,23,45~47]。应用分子辅助标记对胚挽救苗进行早期筛选,能够大大提高育种效率。刘巧[26]和赵雅楠等[21]利用张剑侠等[48]获得的2 个山葡萄抗寒基因连锁RAPD 标记S241-717和S238-854 对胚挽救苗进行早期检测,分别获得携带抗寒分子标记的杂种株系55 个和15 个。
在抗病性方面,少数学者利用分子标记,如抗炭疽病基因RAPD 标记OPC15-1300,抗黑痘病基因RAPD 标记OPS03-1354,抗霜霉病基因RAPD 标记S294 和S382,抗白粉病基因标记ScORN3-R 等,对具有无核基因的胚挽救苗进行了抗病性筛选,获得具有单抗或双抗的无核葡萄新种质。郭海江等[49]得到拥有抗黑痘病基因的胚挽救苗9 株,拥有抗炭疽病基因的胚挽救苗1 株,同时拥有抗炭疽病和抗黑痘病基因的胚挽救苗4 株。张剑侠等[22]得到拥有抗黑痘病基因的杂种4 株。王刚[24]对筛选出的拥有抗霜霉病基因的株系接种霜霉病菌后再次筛选,得到抗病品系10 个。赵炅[25]对65 个胚挽救杂交后代株系进行抗白粉病基因检测,但未扩增出抗白粉病标记的特异性条带。
2 胚挽救技术在无核葡萄育种中取得的成果
我国自20 世纪90年代开始无核葡萄胚挽救研究以来,在胚挽救技术关键因素的优化、三倍体育种、抗性育种等方面进行了大量研究。目前,我国进行无核葡萄胚挽救育种相关研究的主要单位是西北农林科技大学王跃进团队。该团队自1998年起系统开展了无核葡萄胚挽救育种研究,包括对无核葡萄×无核葡萄、无核葡萄×有核葡萄的杂交组合以及无核品种自交的胚挽救操作影响条件的优化,建立了无核葡萄胚挽救技术体系;以中国野生葡萄做父本与欧亚种无核葡萄杂交,进行抗病、抗寒无核葡萄胚挽救育种,获得了一批中国野生葡萄与欧洲葡萄杂交F1代优系,选育出大粒无核抗病优系00-3-1、大粒无核优系00-2-7 等,以及秦红无核、秦翠无核、秦红一号等[4]新品种。
天津师范大学/天津市农业科学院田淑芬团队、北京市农林科学院林业果树研究所徐海英团队、沈阳农业大学、上海市农业科学院、新疆葡萄瓜果开发研究中心、河北省农林科学院昌黎果树研究所等也利用胚挽救技术,在香味、抗病、抗逆无核葡萄育种方面进行了大量研究。其中,河北省农林科学院昌黎果树研究所赵胜建团队较早应用胚挽救技术育成了三倍体葡萄品种无核早红[42];上海市农业科学院蒋爱丽团队先后育成了无核葡萄新品种沪培1 号[50]、沪培2 号[17]、申爱[51]和沪培3 号[18]。
3 无核葡萄胚挽救技术的影响因素
3.1 亲本基因型
母本材料胚的可挽救性是无核葡萄杂交胚能否成功挽救的关键因素[4,9]。现有的研究结果表明,红宝石无核[7,9,26,36]、火焰无核[7,23,31,36]、昆香无核[7,25,31]、奥迪亚无核[52]、京早晶、奥兰多无核、红脸无核、黎明无核、京早白、爱神玫瑰[23]等可挽救性比较好,适宜做母本;无核白、希姆劳特、波尔莱特、优无核、克瑞森无核等胚过早发生败育或可挽救性差的品种不适宜做母本[31,53,54]。唐冬梅[8]对几个无核品种胚挽救的难易程度进行了比较,结果显示,胚挽救由易到难的品种顺序为奥迪亚无核、奇妙无核、火焰无核、优无核、无核紫、皇家秋天、底来特、黎明无核、无核白、森田尼无核。田淑芬等[52]对不同品种自交胚的发育率和萌发率进行了比较,结果显示,奥迪亚无核自交胚的发育率和萌发率最高,其次是红宝石无核、希姆劳特,森田尼无核最低。
杂交父本对胚挽救的成功率也有影响。通常用无核性状传递能力强的品种如无核白、无核黑,或起源于二者的无核品种或品系做父本,后代可分离出较高比例的无核个体[4,53]。郭雨瑞[31]进行了提高无核葡萄胚挽救育种效率的研究,结果显示,玫瑰香和北冰红分别与火焰无核杂交时,利用玫瑰香的挽救效果优于北冰红;当昆香无核做母本时,北冰红做父本的挽救效果优于泰山-2 做父本;当奇妙无核做母本时,阳光玫瑰和双优做父本的挽救效果优于北冰红做父本。史文静等[7]研究指出,香味品种中玫瑰香适合作为父本进行香味无核种质的选育。牛茹萱等[47]报道,欧山杂种较山葡萄更适宜于做杂交父本。
3.2 接种时期
接种时期是影响胚挽救效果的另一关键因素。研究表明,在胚大量败育前、胚发育至心形胚或更高程度时进行胚挽救对于成功挽救具有决定性影响。但迄今为止,还没有一个普遍适用的选择最佳接种时期的方法或指标。目前,大多数学者采用授粉后天数或盛花后天数作为接种时期的具体指标来间接反映胚珠内胚的发育程度。研究表明,不同品种和杂交组合的最佳接种时间不尽相同[7,9,10,31,54~57](表1)。
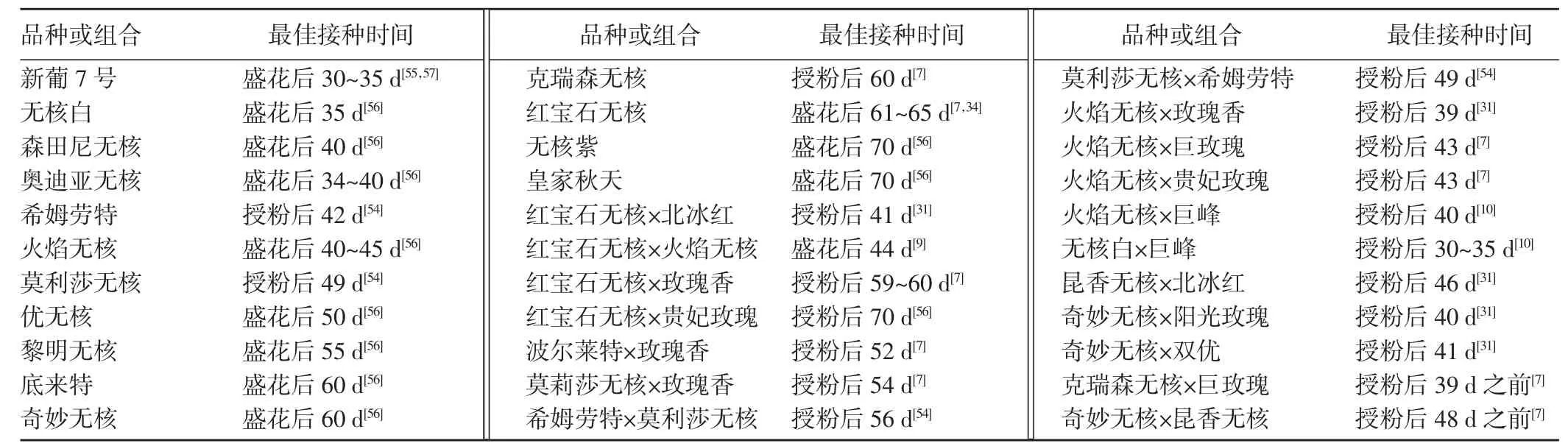
表1 部分葡萄品种和杂交组合的最佳接种时间Table 1 Optimum inoculation time of some grape varieties and hybrid combinations (d)
相同的品种及杂交组合,因植株状况、地理环境或者研究者取样间隔的不同,最佳采样时间会有所差异。田淑芬等[52]研究指出希姆劳特自交胚的最佳取样期为授粉后54 d,而此前报道的希姆劳特自交胚最佳采样时间为授粉后42 d[54]。红宝石无核在南方的最佳采样期为花后33~41 d[32]。父本基因型对杂交组合的取样时间也有一定的影响[24]。实践中,需针对具体的亲本组合进行采样时间的优化。
3.3 培养基
3.3.1 基础培养基 无核葡萄的胚挽救过程分为胚珠内幼胚发育、胚萌发、胚成苗3 个阶段。在胚珠培养阶段应用的培养基有ER、MS、B5、White、Nitsch、NN、改良MM3、MM4 等,其中,Nitsch 培养基和ER 培 养基应 用 得 较 多[29,36,47,57,58]。但 不 同 品种或杂交组合的最适培养基不同,如,波尔莱特做母本时适合ER 培养基,红无籽露做母本时适合MM4 培养基[47]。赵雅楠等[21]对接种于MM3 和ER基本培养基上胚珠的胚发育率和成苗率进行比较后发现,MM3培养基处理的指标值均高于ER 培养基处理。在胚萌发和成苗阶段应用的培养基有Nitsch、ER、BD、1/2 MS、WPM、MM4,其中,使用较多的是1/2 MS 和WPM[7,36,37,47,59,60]。
3.3.2 外源添加物 在外植体培养中,适宜的外源激素种类及配比起着重要作用。不同葡萄品种对激素的敏感程度存在显著差异,并且随胚的发育程度而变化。胚发育程度越高,对激素的要求越不严格。多数研究者认为,培养基中添加适宜浓度的外源激素如6-BA、IAA、IBA、GA3等对胚的发育具有促进作用[3,29,30,36,54]。不同杂交组合对6-BA 的敏感程度不同,其中,应用较多的添加量为0.5 mg/L[29,30,54]。郭雨瑞[31]研究了多效唑(paclobutrazol,PAC)对胚挽救效果的影响,结果显示,在MM3 培养基中添加1.5 mg/L 的PAC 可以提高‘火焰无核×北冰红’、‘昆香无核×泰山-2’杂交胚珠的成胚率;在胚萌发阶段,向WPM培养基中添加1 μmol/L 的PAC 可以提高‘红无籽露×北冰红’的发芽率和成苗率。
在葡萄生长过程中喷施植物生长调节剂也有利于胚珠的发育。唐冬梅[8]研究表明,花前两周喷施100~500 mg/L 的矮壮素能显著促进森田尼无核胚珠的发育,提高森田尼无核胚的萌发率和最终成苗率。李桂荣[60]发现,在无核葡萄坐果过程中喷施IAA 30 mg/L+ABA 30 mg/L 可以促进浆果内胚珠的发育。朱佩佩[36]研究指出,花前2 周喷施30~50 mg/L 的6-BA 能够促进杂交组合胚的发育,这与陶建敏等[61]的研究结果相一致。
添加多胺(腐胺、精胺、亚精胺)[9,60]、氨基酸(丝氨酸、甘氨酸、半胱氨酸、脯氨酸、谷氨酰胺、天冬酰胺)[3,21]、水解酪蛋白、秋水仙碱、麦芽糖、香蕉泥等,也对胚的发育起到一定的促进作用。在胚萌发培养基WPM 中添加3 mmol/L 腐胺有利于‘红无籽露×双优’、‘红宝石无核×紫香无核’、‘爱神玫瑰×阳光玫瑰’成苗,成苗率分别提高了13.34%、6.67%和30.16%;添加0.5 mmol/L 亚精胺有利于‘红宝石无核×火焰无核’、‘红脸无核×爱神玫瑰’成苗;添加0.3 mmol/L 精胺有利于‘红宝石无核×江西-2’、‘红无籽露×紫香无核’成苗[9]。田淑芬等[3]研究了不同甘氨酸浓度对胚发育的影响,认为不同杂交组合使用的最佳甘氨酸浓度也不一样,其中,‘红宝石无核×玫瑰香优系’使用的最适浓度为4.0 mmol/L,‘希姆劳特×玫瑰香优系’使用的最适浓度为1.0 mmol/L。
3.3.3 培养基形态 胚挽救过程中培养基的形态主要有固相、液相和固液双相3 种。其中,固液双相培养基弥补了单相培养基的缺陷,既能增大胚珠与培养基的接触面积,又能避免培养基因水分蒸发过度而导致的浓缩,经检验[32,36,55,56,60]培养效果很好。但也有研究显示,对于胚珠较大的品种,3 种相态培养基的挽救效果差异不大[56]。
3.4 培养条件
培养时的光照、温度、珠被处理方式等对无核葡萄胚挽救也有一定的影响。
无核葡萄胚挽救时胚珠培养过程中是否需要光照,目前研究结果不完全一致。由于胚在胚珠内的发育是不见光的,于是,唐冬梅等[56]和李桂荣[60]模拟胚在胚珠中的环境状态,比较了暗培养和光照培养(光照与黑暗交替)对胚发育的影响,研究结果均证实暗培养有利于离体胚的发育。史文静等[7]认为,暗培养时间在8~10 周为宜,暗培养时间过长会显著增加畸形胚的比例。但郑婷[32]的研究结果显示,光照(光照与黑暗交替)培养可以促进胚的发育。李玉玲等[27]探讨了不同光质(红光、蓝光、白光、暗)培养对火焰无核胚珠培养的影响,发现4 种光质处理对胚发育的影响效果差异不显著。
对于胚培养过程中是否都需要低温,研究者的看法也不一样。一般认为,为打破胚的休眠,促进胚萌发,提高萌发率和培养壮苗,需对胚珠进行一定时间的4 ℃低温处理。郑婷[32]研究表明,低温处理‘火焰无核×寒香蜜’胚珠10 d,可以使胚珠发育率、胚发育率和胚萌发率均达到相对较高的水平。但也有学者认为,由于无核葡萄的胚未发育成熟,其并未进入休眠状态,因此,无需考虑休眠问题。唐冬梅等[56]和田淑芬等[52]的研究结果均表明,在胚珠离体培养阶段低温培养会抑制胚的发育,降低胚发育率。
在无核葡萄胚挽救过程中,珠被的处理方式有不处理(全胚珠)、横切、纵切、切喙和剥取裸胚。多数学者认为,剥取裸胚可以提高胚的萌发率[62],使胚提前萌发且成苗整齐,同时,可直接判断胚挽救苗是来自合子胚还是来自体细胞胚。但是,由于不同的品种或杂交组合胚剥离难易程度不一样,且在剥胚过程中容易使胚受伤而产生较多的畸形苗,故一些学者采取横切[63]、切喙[58]的方式处理珠被,使胚在胚乳和培养基的双重营养下萌发生长,同样可以提高胚珠的萌发出苗率,并且操作容易,对胚的伤害小。所以,横切和切喙的珠被处理方式也被较多学者所应用。
3.5 胚挽救苗的驯化移栽
胚挽救苗的驯化移栽是一个由半异养到自养、高湿到低湿、恒温到变温、无菌到有菌的过程[33,64],是其从实验室走向大田的关键步骤。李桂荣[60]建立的移栽驯化操作程序为强光闭口炼苗7~14 d—移栽至基质—罩透明塑料杯保湿炼苗—温室驯化—移植大田。胚挽救苗的生长状况、移栽基质和喷施营养液等是影响无核葡萄育种成苗的主要因素。
一般认为,苗龄30~40 d 是胚挽救苗移栽的适宜时期[6,8,33,64];60 d 时的试管苗根系已经老化,移栽成活率会大大降低。但是,苗龄并不适合所有的情况,应结合要移栽驯化的胚挽救苗的长势和高度进行综合评判。李桂荣[60]研究认为,苗高8~10 cm 时是胚挽救苗的最适移栽时期。
单一的移栽基质成分有园土、泥炭土、蛭石、河沙、珍珠岩、椰糠等。一般认为,应用复配基质的效果优于应用单种特定的移栽基质。实践中应用的复配基质配方有V(珍珠岩)∶V(蛭石)∶V(河沙)=1∶1∶1[33]、V(珍珠岩)∶V(草炭)∶V(园土)=4∶1∶1[6]、V(珍珠岩)∶V(草灰)=1∶1[62]、V(基质土)∶V(蛭石)=5∶1[7]、V(蛭石)∶V(泥炭土)∶V(椰糠)=1∶4∶1[60],V(椰糠):V(蛭石)∶V(腐叶土)=2∶1∶1[65]。驯化移栽时常用的营养液为B5 和MS。对营养液浓度进行比较后,多数学者认为,用1/8 MS大量元素[8,24,33,60]营养液浇灌胚挽救苗利于壮苗,有助于移栽成活率的提高;也有人认为,应用更低浓度的1/10 MS[60]或1/16 MS[6]营养液对胚挽救苗的移栽驯化有利且经济。唐冬梅[8]用1/4 B5 大量元素溶液浇灌移栽苗,幼苗的成活和生长效果较好。
生根粉属于生长素类的植物生长调节剂,生长素能调控侧根根原基的形成,促进植物侧根和不定根的发生与生长,并且侧根的发生与伸长都直接或间接地依赖于生长素的调控作用[59]。用300 mg/L 生根粉浸根20~30 s[33]或用1.0 mg/L 生根粉浸根3 min[59]处理幼苗,移栽成活率较高。
4 展望
我国在葡萄育种中应用胚挽救技术已有30 余年的历史,在无核葡萄胚挽救影响因素方面以及三倍体育种、远缘杂交育种和抗性育种等方面进行了系列研究,建立了无核葡萄胚挽救技术体系,创制了很多无核葡萄新种质,并育成一些无核葡萄新品种。但与国外引进品种相比,我国的这些育成品种在商品性上缺乏突破;且多数品种都是由2 个外来或古老种质的简单杂交获得,缺少多亲多代综合杂交育成的品种[66]。这一状况与我国拥有众多栽培品种和丰富野生葡萄资源的事实不相符。我国野生葡萄中的一些种和株系具有较强的抗病、抗逆性,现有研究表明其基因组存在主效抗病基因,且与欧洲葡萄无核品种杂交易亲和,因此,在对葡萄重要性状的遗传规律进行系统研究的基础上,充分利用我国的葡萄资源,应用胚挽救技术与传统杂交育种等手段,选育具有大粒、无核、抗性强等优良性状的葡萄新品种仍是目前育种工作者不断探索和研究的重点。
利用胚挽救技术得到的F1代杂种中还存在一定比例的有核杂种,尤其是利用有核葡萄做父本时。利用分子标记对F1代杂种进行无核、抗性性状的早期筛选,能够提高育种效率,节省大量人力、物力、财力。迄今,已有5 个无核基因分子标记被用于葡萄无核性状的辅助选择,分别是SCAR 标记SCC8、SCF27 和GSLP1-569,微卫星标记VMC7F2 和p3-VvAGL11[4,67]。在抗病性方面应用的分子标记有许多,如,抗炭疽病基因PAPD 标记OPC15-1300,抗黑痘病基因RAPD 标记OPS03-1354,抗霜霉病基因RAPD 标记S294 和S382,抗白粉病基因RAPD 标记OPW02-1756 等。在抗寒性方面应用的分子标记有S241-717 和S238-854。由于分子标记与目标基因的连锁程度、适用的广泛性等影响着辅助选择的效率,因此,继续寻找紧密连锁的无核基因分子标记以及抗病和抗逆基因分子标记,仍是今后葡萄分子育种的重要内容。
