细粒棘球蚴重组抗原蛋白BCG-egG1Y162免疫原性的初步研究
2019-05-31李海丽刘永太张峰波李玉娇李智伟丁剑冰王红英
李海丽, 刘永太, 张峰波, 李玉娇, 李智伟, 赵 骁, 丁剑冰,4, 王红英
(新疆医科大学1第一临床医学院, 2基础医学院, 乌鲁木齐 830011; 3阿克苏市兵团第一师医院检验科, 新疆 阿克苏 843000;4省部共建中亚高发病成因与防治国家重点实验室, 乌鲁木齐 830011)
细粒棘球蚴病(cystic echinoccosis, CE)是一种由细粒棘球绦虫(Echinococcusgranulosus, Eg)幼虫引起的人兽共患病,随着世界畜牧业的发展不断扩散[1]该病呈全球分布,而中国更是CE高发的国家之一,主要分布在新疆北部、青海以及宁夏等地区[2]。细粒棘球蚴病在早期不易被发现,潜伏期长,发展到后期会给人类生产、生活造成极大危害[3]。目前包虫囊肿的主流治疗方案是通过外科手术摘除囊肿,不宜进行手术的患者则通过大剂量甲苯咪唑控制囊肿生长[4]。然而,包虫病的术后复发率高且治疗作用有限,所以疫苗预防是细粒棘球蚴病最经济有效的治疗方法[5]。
多年来国内外学者在细粒棘球蚴病的免疫学和疫苗研制方面开展了大量工作。Lightowlers等[6]在1993年研制出了抗细粒棘球绦虫虫卵感染的基因工程重组抗原疫苗,命名为Eg95。Eg95是目前应用最广泛的抗细粒棘球蚴病的疫苗,在新西兰、澳大利亚和阿根廷的预防效果较好,但是在亚洲中部和中国西北部地区并不能发挥理想的作用[7-8]。因此,科研工作者希望能够开发出高效且特异性高的疫苗来抵抗细粒棘球蚴病。
前期研究发现细粒棘球蚴病抗原egG1Y162的基因序列、氨基酸序列与蛋白结构都与多房棘球蚴的候选疫苗EmY162存在高度相似的情况[9]。基于重组抗原EmY162对终末宿主的良好保护作用,以及重组卡介苗(BCG)活体疫苗载体的特点,本实验室在egG1Y162抗原的基础上成功构建了rBCG-egG1Y162菌株,即将egG1Y162抗原基因克隆到大肠埃希菌-分枝杆菌的穿梭表达载体pMV361中,再通过电转法将携带目的基因的质粒转入BCG中,从而诱导表达BCG-egG1Y162重组蛋白[10-11]。在此基础上,希望通过研究重组抗原蛋白BCG-egG1Y162免疫小鼠后对机体免疫力的影响,评价小鼠的相关体液免疫、细胞免疫指标,以及抗细粒棘球蚴感染的能力,来为CE的防治提供理论依据。
1 材料和方法
1.1 材料
1.1.1 实验动物 6~8周BALB/c雌性小鼠60只,体质量18~22 g,由新疆医科大学第一附属医院临床研究院实验动物科学研究部提供。
1.1.2 细粒棘球蚴原头蚴 从新疆医科大学第一附属医院肝胆外科包病虫患者手术摘除的完整包囊中无菌条件抽取囊液,分离原头蚴,用生理盐水配制成1.5×104个原头蚴/mL,用于感染BALB/c雌性小鼠。
1.1.3 细粒棘球重组蛋白BCG-egG1Y162 由本室提供。将egG1Y162抗原基因克隆到大肠埃希菌-分枝杆菌的穿梭表达载体pMV361中,再通过电转法将携带目的基因的质粒转入BCG中,从而诱导表达BCG-egG1Y162重组蛋白[12]。
1.1.4 主要试剂 大肠埃希菌-分支杆菌穿梭质粒pMV361和BCG由石河子大学医学院张万江教授惠赠;辣根过氧化物酶标记的羊抗鼠IgG、刀豆蛋白A (ConA)、四甲基联苯胺(TMB)、四甲基偶氮唑蓝(MTT)购买自华美生物工程公司;IL-4、IFN-γ的ELISA检测试剂盒购买自福麦斯公司;弗氏完全佐剂和不完全佐剂(IFA)购买自Sigma公司;RPMI1640培养基购买自Gibico公司。
1.2 方法
1.2.1 动物分组及免疫接种 随机将BALB/c雌性小鼠分成3组,每组20只,A组为BCG-egG1Y162免疫组,B组为重组卡介苗组(BCG组),C组为正常组(生理盐水对照组)。A组即免疫组小鼠,在小鼠背部多点皮下注射200 μL重组蛋白BCG-egG1Y162(25 μg/μL,重组蛋白用PBS溶解后加等体积FCA乳化),先后共免疫4次,后3次加强免疫时间间隔为2周;B组为BCG组,小鼠背部多点注射100 μL BCG+100 μL FCA,免疫次数和间隔时间同A组。A、B2组首次免疫用完全弗氏佐剂,后3次加强免疫用不完全弗氏佐剂;C组为正常组,背部多点皮下注射生理盐水200 μL /只/次。在免疫的第0、2、4、6、8、32周通过毛细管进行眼底静脉丛取血,离心机预冷至4℃,外周血2 000 r/min离心10 min获得血清,置于-80℃冰箱中保存。在免疫的第8周,每组随机处死10只小鼠用于分离脾淋巴细胞,剩下的10只小鼠用于体内抗活棘球蚴原头蚴感染实验。
1.2.2 ELISA法检测血清中IgG的水平 采用ELISA法检测各组0、2、4、6、8、32周的血清中IgG抗体水平的动态变化。用包被液稀释BCG-egG1Y162调整浓度至10 μg/mL,按100 μL加入到酶标板各孔中,置于4℃层析柜中包埋过夜。次日用PBS-0.5%Tween洗板3次,5%脱脂奶粉封闭2 h封闭非特异性位点。弃去封闭液后重复洗板步骤,再加入按1:200的稀释比稀释各组血清,100 μL /孔37℃孵育2 h,每个样本设置3个复孔。重复洗板3次后,加入1∶1 000稀释的羊抗鼠IgG-HRP 100 μL/孔,37℃孵育2 h。PBS-0.5%Tween洗板3次,加入TMB显色15 min,用50 μL /孔的H2SO4终止反应,酶标仪读取450 nm处各孔吸光度(OD值)。
1.2.3 脾细胞悬液的制备 在免疫的第8周每组随机处死10只BALB/c小鼠,置75%乙醇中消毒5 min。超净台中解剖取脾脏,匀浆,用2 mL PBS冲洗,200目尼龙膜过筛,滤液加入10倍体积的红细胞裂解液,2 000 r/min 离心5 min。PBS洗3次,2 000 r/min 离心5 min,弃去上清。用RPMI1640调整密度至5×106/mL。台盼蓝染色显示脾细胞存活率达到95%以上,可用于后续实验。
1.2.4 BCG-egG1Y162刺激后脾淋巴细胞上清中IFN-γ、IL-4水平的测定 将浓度为5×106/mL的各组小鼠的脾淋巴细胞悬液分别加入到96孔细胞培养板中,加入10 μg/mL的BCG-egG1Y162重组抗原蛋白刺激72 h,以未加刺激因子的脾淋巴细胞组为空白对照,均重复6孔,细胞培养板置于37℃、5%CO2恒温培养箱中常规培养。结束后收集脾细胞悬液,1 000 r/min离心15 min获取细胞上清液,用IFN-γ、IL-4试剂盒分别检测小鼠脾细胞培养上清中的IFN-γ、IL-4的含量水平,来更进一步的探讨BCG-egG1Y162诱导的免疫反应中是细胞免疫还是体液免疫发挥主要作用。
1.2.5 小鼠脾淋巴细胞的增殖实验 将调整好浓度的各组小鼠的脾淋巴细胞悬液加入到96孔细胞培养板中,分别用5 μg/mL ConA以及10 μg/mL BCG-egG1Y162刺激72 h,以未加刺激因子的脾淋巴细胞组为空白对照,均设置6个复孔,置于37℃,5%CO2条件下培养。培养结束前4 h,加入20 μL MTT(5mg/mL),继续培养至规定时间。结束后2 000 r/min离心8 min,弃去上清,加入100 μL二甲基亚砜低温振荡10 min,轻轻反复吹打至甲簪充分溶解,测定样本在570 nm处的吸光度(OD值)。
1.2.6 活棘球蚴原头蚴感染小鼠和免疫保护力的测定 在第8周时,A、B、C3组小鼠腹腔注射活棘球蚴原头蚴1500枚/只,在第32周(感染后24周)处死小鼠,打开腹腔,对各组小鼠腹腔内包囊个数进行计数并测量包囊直径,根据公式计算出各组小鼠的免疫保护力,免疫保护力(%)=(1-实验组平均包囊数/正常组平均包囊数)×100%。
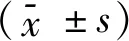
2 结果
2.1 BCG-egG1Y162免疫小鼠后血清中IgG动态观察ELISA法检测各组小鼠血清中抗体IgG在第0、2、4、6、8周的OD值水平,结果如表1所示。BCG-egG1Y162免疫组血清中IgG抗体从第4周开始表现为阳性,并且持续增长,在第8周达到最高水平。经分析,BCG-egG1Y162免疫组IgG水平从第4周开始就显著高于第0周,差异具有统计学意义(P<0.05);从第4周直到第8周,BCG-egG1Y162免疫组的IgG水平与同一时间的BCG组和正常组相比,差异具有统计学意义(P<0.05);BCG组和正常组之间差异无统计学意义(P>0.05),结果见表1。

表1 BCG-egG1Y162免疫小鼠后血清中IgG动态观察(OD Value, ±s)
注: 与正常组或BCG组相比,*P<0.05。
2.2 小鼠脾淋巴细胞增殖能力检测结果将A、B、C3组小鼠脾淋巴细胞分别与ConA、重组蛋白BCG-egG1Y162共同培养,另外设置未加刺激物的组别作为对照,MTT法检测3种培养条件下的各组小鼠脾淋巴细胞的增殖情况。结果发现,ConA刺激可使3组小鼠脾淋巴细胞的增殖能力得到明显提高,与同组未加刺激物相比差异具有统计学意义(P<0.05),表明各组小鼠脾淋巴细胞在ConA的刺激下均有增殖能力。但BCG-egG1Y162免疫组、正常组、BCG组3组之间差异均无统计学意义(P>0.05)。用重组蛋白BCG-egG1Y162刺激脾淋巴细胞后,BCG-egG1Y162免疫组脾淋巴细胞与BCG组和正常组相比明显增加(P<0.05),与同组无刺激物相比差异具有统计学意义(表2)。并且BCG组和正常组之间差异无统计学意义(P>0.05)。提示BCG-egG1Y162免疫组的脾淋巴细胞在体外能被BCG-egG1Y162蛋白诱导产生较好的免疫记忆能力,能够被特异性的刺激增生,诱生一定水平的细胞免疫。
2.3 小鼠脾淋巴细胞上清中IFN-γ、IL-4的诱生和测定结果在BCG-egG1Y162免疫小鼠的脾淋巴细胞上清液中,IFN-γ、IL-4的表达量以及IFN-γ/IL-4的比值显著高于正常组和BCG组(P<0.05)。IFN-γ、IL-4的表达量及其IFN-γ/IL-4比值在正常组和BCG组中无统计学差异(P>0.05),见表3。由此可见,重组蛋白BCG-egG1Y162具有免疫原性,并且显著增强了小鼠的脾淋巴细胞的Th1/Th2免疫反应,平衡偏向Th1反应,即细胞免疫占主导地位。
表2 免疫小鼠脾淋巴细胞增殖水平(OD Value, ±s)
注: 与正常组或BCG组相比,*P<0.05。

表3 小鼠脾淋巴细胞上清中IFN-γ、IL-4的水平及其比值
注: 与正常组或BCG组相比,*P<0.05。
2.4 小鼠感染活棘球蚴原头蚴后血清中IgG水平检测ELISA法检测各组小鼠感染活棘球蚴原头蚴后血清中的IgG的OD值水平,如表4所示。BCG-egG1Y162免疫组在感染活棘球蚴原头蚴后的第24周(免疫后第32周)时,小鼠血清中的IgG水平相对于感染第8周有所下降,与第0周相比差异有统计学意义(P<0.05),并且显著高于同一时间的BCG组和正常组(P<0.05)。BCG组和正常组在感染后第24周的小鼠血清中的IgG水平与第0周和第8周相比差异无统计学意义(P>0.05),2组在同一时间的IgG水平差异无统计学意义(P>0.05)。
表4 BCG-egG1Y162免疫小鼠感染活棘球蚴原头蚴后血清中IgG水平(OD Value, ±s)
注: 与正常组或BCG组相比,*P<0.05。
2.5 小鼠的抗活棘球蚴原头蚴感染能力检测结果A、B、C3组小鼠腹腔内注射活棘球蚴原头蚴1 500枚/只,在感染后第24周(免疫后第32周)处死,计数各组小鼠包囊个数以及直径。发现BCG-egG1Y162免疫小鼠体内的包囊数及包囊大小与BCG组和正常组相比明显减少和缩小,差异具有统计学意义(P<0.05),BCG组和正常组相比无统计学意义(P>0.05)。相比于正常组,发现BCG-egG1Y162可诱导小鼠产生87.1%的免疫保护力抵抗活棘球蚴原头蚴的感染,BCG组的免疫保护力为8.02%,见表5。
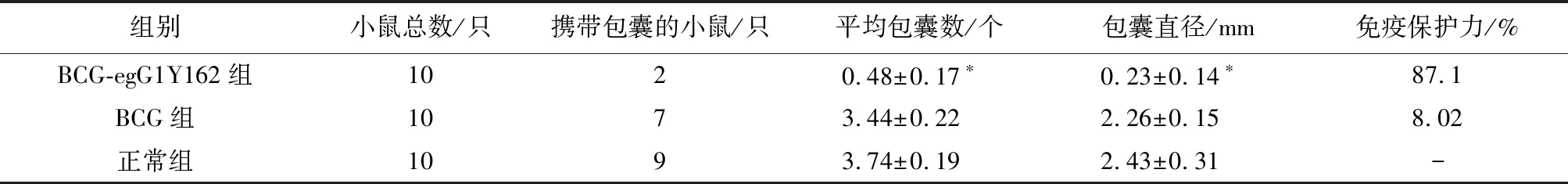
表5 各组小鼠产生的包囊数及免疫保护力
注: 与正常组或BCG组相比,*P<0.05。
3 讨论
细粒棘球蚴病是一种严重危害人类健康的人兽共患寄生虫病,是当前国家卫生部规划防治的重要寄生虫病之一。近年来随着人口流动的增加以及畜牧业的发展,细粒棘球蚴病不断地蔓延和扩散[13]。因此,提高防治工作的力度刻不容缓。相对于外科手术以及药物治疗带来的高复发率和副作用,抗原疫苗的出现为CE的防治开辟了一条新途径。抗原疫苗通过加强机体自身的免疫应答来抵抗细粒棘球蚴病,高效且具有特异性,值得进一步探究[14]。本实验希望通过研究重组蛋白BCG-egG1Y162对机体免疫应答发挥的作用,来为细粒棘球蚴病的防治工作提供一定帮助。
目前一些研究表明体液免疫和细胞免疫在机体抵抗细粒棘球蚴感染中均发挥了重要作用[15]。在体液免疫方面,IgG是全身性体液免疫分子的效应因子,在抗感染中发挥中和毒素以及调理等作用,所以能在一定程度上反映了机体体液免疫的水平[16]。BCG-egG1Y162免疫组小鼠血清中IgG抗体水平从第2周开始逐渐上升,在第8周达到最高水平,从第4周开始其IgG水平与正常组和BCG组相比差异具有统计学意义。表明BCG-egG1Y162可诱导小鼠产生特异性的体液免疫应答,说明BCG-egG1Y162具有较好的免疫原性。
而在细胞免疫中,T淋巴细胞是机体非常关键的活性免疫细胞之一,T淋巴细胞的增殖和分化是免疫应答过程中的一个重要阶段,通过检测脾T淋巴细胞的增殖水平来反映其功能的高低是评价机体细胞免疫功能的常用方法[17]。通过MTT法检测发现,BCG-egG1Y162免疫组小鼠的脾淋巴细胞在重组蛋白BCG-egG1Y162的刺激下,其数目远高于BCG组和正常组,差异具有统计学意义。而在ConA的刺激下,免疫组、正常组和BCG组的脾淋巴细胞虽然均表现出明显增殖,但是3组之间差异无统计学意义。由此推测BCG-egG1Y162免疫可诱导小鼠脾细胞产生较好的记忆能力,在体外能被重组蛋白BCG-egG1Y162特异性刺激增生从而产生细胞免疫应答。
BCG-egG1Y162可诱导机体的体液和细胞免疫应答,为了进一步探究在是何种应答方式在BCG-egG1Y162诱导的抗细粒棘球蚴感染的过程中占主导地位,本研究通过ELISA法检测BCG-egG1Y162免疫组小鼠的脾淋巴细胞上清中IFN-γ、IL-4的水平并比较两者的比值。其中IFN-γ主要由Th1细胞分泌产生,介导细胞免疫。而IL-4由Th2细胞产生,促进体液免疫。有研究表明,IFN-γ促进Th0向Th1分化,增强细胞免疫应答使体液免疫应答降低[18]。IL-4则是促进Th0向Th2分化,增强体液免疫应答的同时对细胞免疫起下调作用[19]。郭忠帅等[20]研究发现,CE患者术后血清中IFN-γ表达上调,是因为随着包囊的切除,相应抗原的减少,宿主体内免疫应答逐渐恢复,机体通过分泌IFN-γ来抵抗细粒棘球蚴感染。所以IFN-γ/IL-4比值反映了Th1/Th2的平衡偏向,即免疫应答中是体液免疫还是细胞免疫占据优势。本研究结果显示,IFN-γ、IL-4以及IFN/IL-4比值均显著性高于正常组和BCG组,说明在BCG-egG1Y162诱导的免疫应答中,细胞免疫发挥主要作用。
在BCG-egG1Y162诱导的小鼠体内抗细粒棘球蚴感染实验中,BCG-egG1Y162免疫组的小鼠在感染细粒棘球蚴后IgG水平显著高于正常组和BCG组,说明BCG-egG1Y162免疫小鼠能增强其抗细粒棘球蚴感染能力。为了进一步探究BCG-egG1Y162的抗感染能力,本研究计数感染后24周的各组小鼠的包囊个数,并比较包囊直径大小,结果发现,BCG-egG1Y162免疫组的小鼠的包囊数目和大小与正常组和BCG组相比较明显减少和缩小,具有统计学意义。以正常组为阴性对照,BCG-egG1Y162免疫组小鼠的免疫保护力达到了87.1%,明显高于BCG组的8.02%,所以BCG-egG1Y162能诱导小鼠抗细粒棘球蚴感染,可成为细粒棘球蚴病的候选疫苗。
综上所述,细粒棘球蚴重组抗原蛋白BCG-egG1Y162免疫小鼠可上调小鼠血清中IgG的水平并刺激脾淋巴细胞增殖活化,特异性的增强脾淋巴细胞的Th1免疫反应,减少感染小鼠包囊的发生。所以重组蛋白能够有效地诱导特异性免疫应答的发生,通过增强体液免疫和细胞免疫发挥抗细粒棘球蚴感染的作用,其中细胞免疫应答发挥主要作用。说明BCG-egG1Y162是具有研究价值的细粒棘球蚴病候选疫苗分子,能为细粒棘球蚴病的防治工作带来帮助。
