双花木属植物潜在分布区模拟与分析
2019-05-31孟艺宏姜小龙徐刚标
孟艺宏,徐 璕,姜小龙,3,徐刚标,*
1 中南林业科技大学林木遗传育种实验室, 长沙 410004 2 四川大学生命科学学院, 成都 610064 3 中国科学院上海辰山植物园, 上海 201602
现代物种地理分布格局受末次冰期和间冰期交替引起的气候变化影响很大,陆生物种部分灭绝,部分在原地通过适应性进化、或迁移至避难所以适应冰期的恶劣气候。幸存下来的陆生植物,在冰期后变暖的气候条件下从避难所扩散、迁移后重新分布[1]。了解物种对历史气候变迁的响应,有助于理解物种分化和形成的历史成因及当今全球气候变暖对物种将来分布格局的影响[2-3],可为物种保育策略的制定提供科学指导,在理论研究和生产实践上都具极其重要的意义[4]。
物种分布模型(Species Distribution Models, SDMs)是以生态位理论为基础,利用物种已知居群分布点及其相关联的气候因子数据,基于一定的算法构建模型,推算物种的生态需求,模拟物种在不同时期的分布区[5-6]。目前,物种分布模型有多种,包括分类与回归树(Classification And Regression Tree, CART)、最大熵(Maximum Entropy, MaxEnt)、基于规则集的遗传算法(Genetic Algorithm for Rule-set Prediction, GARP)等模型。MaxEnt 模型是以研究地理区域的所有栅格构成最大熵的可能分布空间,以物种当前已知分布栅格的环境变量(连续型和分类型)作为约束条件,寻找约束条件下最大熵的概率分布作为最优分布来预测物种潜在分布区[7]。MaxEnt 模型具有建模简单、预测效果精确、结果易于解释等优点[8- 10],已被广泛应用于濒危物种保护[11]、外来物种入侵[12]、全球气候变化对物种分布的影响[13]和物种谱系地理[14- 15]等领域研究。
双花木属(DisanthusMaxim.)隶属金缕梅科(Hamamelidaceae),为东亚地区特有属,1种1变种,被世界自然保护联盟(http://www.iucn.org/)列为濒危物种,在研究金缕梅科系统发育和东亚植物区系地理演化等方面具有重要的科学价值。该属植物的叶片及花粉化石发现于欧洲及格鲁吉亚的中新世,种子化石发现于中欧地区白垩纪最晚期[16]。目前,双花木属植物居群小,数量少。原种双花木(D.cercidifoliusMaxim.)仅残存于日本南部本州和四国山地[17],变种长柄双花木(D.cercidifoliusMaxim. var.longipesH. T. Chang)间断分布于我国南岭地区、武夷山脉海拔1300 m以下的山区[18]。
双花木属植物为落叶小乔木或灌木,叶心型,花果红色,为观赏性高的珍稀濒危植物[19]。目前,双花木属植物的研究主要集中于资源分布[18]、系统发育[17,20]、遗传多样性[21-22]和群落生态[23]等方面,潜在地理分布区模拟尚未见报道。因此,有关双花木属植物地理分布与气候之间关系、过去与未来不同气候情景下适生区等居群进化信息尚不清楚。本研究采用MaxEnt 3.2.19和ArcGIS 10.0软件,模拟双花木属植物在不同历史时期的地理分布动态,探讨末次盛冰期、当前及未来不同气候情景下该属植物适生分布范围,以期重建双花木属进化过程中地理分布的变迁历史,探讨限制该属植物地理分布的主导气候变量,为双花木属植物保育措施的制定和东亚植物区系物种形成演化的研究提供理论依据。
1 材料与方法
1.1 双花木属地理分布数据获得及处理
考虑到居群分布点的气候数据间存在空间自相关性,可能导致分布区模拟结果产生偏差,地图上2.5′×2.5′的网格中仅保留1个点作为有效居群分布点。本研究共筛选出19个双花木属植物居群分布点,其中13个长柄双花木居群地理数据源于野外实地调查,6个双花木居群地理数据源于文献记载[17]。
1.2 气候变量筛选
从WorldClim数据库(http://worldclim.org)中,分别下载空间分辨率为2.5′的末次盛冰期(约22000年前)、当前(1950—2000年)、未来(2060—2080年)气候数据。未来气候数据包括辐射强度分别为2.6、4.5、6.0、8.5 W/m2的4种温室气体排放情景,其典型浓度路径(Representative Concentration Pathways, RCPs)分别为2.6、4.5、6.0和8.5。每个时期的气候数据包括与月降水量、月均温、月最高温、月最低温相关的19个气候变量。
Maxent模型对选取的居群地理数据敏感[7],气候变量间相关性过高容易产生过度拟合[24]。因此,采用ArcGIS10.0提取双花木属植物各居群的气候变量信息,运用R语言中Dismo package(http://CRAN.R -project. org/package=dismo),检验气候变量间的相关性。基于相关性大于0.95的1对气候变量中只选1个变量的原则选取气候变量,用于Maxent模型运算。
1.3 模型准确性评价
由于双花木属植物居群数量少,采用留一法模拟潜在分布范围。依次去掉19个居群中的一个居群,利用剩余的18个居群用于Maxent模型模拟,共模拟19次。基于ArcGIS 10.0软件,将19次模拟结果的平均值进行分布区的气候适生等级划分和可视化处理。分布区的适生等级分为低适(0.2—0.4)、较适(0.4—0.6)、适生(0.6—0.8)和最适(0.8—1.0)4个级别。将不同居群的地理位置导入预测图,基于预测图等级划分与当前分布区的一致性,检验模拟结果的可靠性。
采用MaxEnt 3.2.19软件建模,制作受试者工作特征曲线(receiver operating characteristic curve,ROC)。以ROC曲线下方面积 (area under receiver operating characteristic curve,AUC)值,评价模型的有效性。AUC取值范围为[0.5, 1]。AUC值0.5—0.7,模拟效果差;0.7—0.8,模拟效果一般;0.8—0.9,效果较好;0.9—1.0,效果极好[25]。
1.4 气候变量重要性评估
运行MaxEnt 3.2.19软件过程中,勾选“do jackknife to measure variable importance”,基于刀切法(jackknife),计算各气候变量的测试增益。结合MaxEnt 3.2.19软件自动生成变量贡献率(percent contribution),推测限制物种分布的主导气候变量。
1.5 不同时期分布区变化估算
选择MaxEnt 3.2.19软件模拟结果中的最大训练敏感度和特异性阈值(maxinum training sensitivity plus specificity threshold)作为物种存在/不存在的阈值[26],运行ArcGIS 10.0软件,生成各个时期的物种分布图,采用ArcToolbox中的提取工具提取各个时期居群生境的适宜性数值,将不同时期的双花木属分布图进行叠加,统计各时期分布区及生态位重叠的栅格数量。基于生境稳定性(Nstab)、当前分布面积与其他时期分布面积的比值(Na)和扩张或收缩程度(Ne),比较双花木属在不同时期的地理分布变化[27]。
Nstab=1-|Npre-Nx|
式中,Npre和Nx分别代表当前和其他时期(LGM、RCP 2.6、RCP 4.5、RCP 6.0和RCP 8.5)居群生境适宜性,居群生境适宜性划分为高(0.8—1.0)、中(0.6—0.8)、低(<0.6),Nstab表示居群生境稳定性,居群生境稳定性等级划分为高(0.9—1.0)、中(0.8—0.9)、低(<0.8)。Na=当前分布区面积/其他时期分布区面积,Ne=1-(其他时期和当前生态位重叠区域面积/当前分布区面积)×100%,Ne表示物种的扩张或收缩程度。
2 研究结果
2.1 MaxEnt模型准确性
ROC曲线检验结果表明,各个时期19次重复模拟的测试集和训练集AUC的平均值均达0.999,标准差为0.0001,几乎接近于1。这说明,MaxEnt模型模拟双花木属植物在不同时期地理分布区的效果很好,可信度高,模拟的双花木属植物地理分布范围与当前自然分布区高度吻合。
2.2 双花木属植物分布区气候适宜性

图1 不同时期的双花木属潜在分布格局Fig.1 The potential distribution patterns inDisanthusMaxim. in different periods红色圆点代表双花木属居群位置,红色圆点的大小表示生境适宜性的高低
MaxEnt 3.2.19模拟的结果经ArcGIS 10.0软件可视化处理的结果(图1)显示,不同时期,双花木属植物的潜在地理分布范围存在差异。末次盛冰期(图1A),双花木属植物最适分布区为日本宫崎、千叶地区,适生分布区为中国南岭山脉、日本九州和中部地区以及露出海平面的东海大陆架。当前(图1B),双花木属植物最适分布区为中国南岭、武夷山东麓与黄山、罗霄山山脉,以及日本纪伊山、赤石山及阿尔卑斯山脉,适生分布区为中国台湾雪山、朝鲜太白山等局部地区。未来,随着温室气体排放量升高,双花木属植物适生分布面积逐渐减小(图1C—F)。气候变化对日本双花木分布区的影响远小于对中国长柄双花木分布区的影响,中国将不再有长柄双花木最适宜的分布区。RCP 2.6(图1C)与RCP 4.5(图1D)气候情景下,长柄双花木适生区为南岭、黄山和武夷山东麓的部分地区以及罗霄山山脉的局部地区;双花木适生分布范围变化不大,仅日本九州适生分布区面积减少。RCP 6.0气候情景下(图1E),中国南岭山脉长柄双花木适生分布面积减少,但日本九州地区气候条件更适宜双花木生长。RCP 8.5气候场景下(图1F),中国武夷山山脉气候不适宜长柄双花木生长,日本九州地区生境破碎化且双花木分布的适生性大大下降。
2.3 双花木属植物居群生境适宜性
采用ArcToolbox中的提取工具提取不同时期各居群生境适应性值的结果显示,末次盛冰期(图1A),中国邵阳、宜黄、龙泉及日本广岛、滋贺、长野、歧阜居群生境适宜性低,其他居群生境适宜性中等。当前(图1B),仅中国井冈山居群生境适宜性低,中国莽山、邵阳、蓝山、连州及日本冈山和滋贺居群生境适宜性中等,其他居群生境适宜性高。未来,RCP 2.6气候情景下(图1C),中国莽山、蓝山、宜黄、井冈山、连州及日本冈山居群生境适宜性低;RCP 4.5气候情景(图1D)下,中国莽山、蓝山、井冈山、连州及日本冈山、广岛、滋贺居群生境适宜性低;RCP 6.0气候情景(图1E)下,中国莽山、蓝山、井冈山、连州、道县、宜章、新宁、邵阳居群生境适宜性低;RCP 8.5气候情景(图1F),中国道县、莽山、蓝山、宜章、连州、新宁、邵阳、宜黄、宜丰、井冈山、龙泉、开化、日本广岛、滋贺居群生境适宜性低。总体来看,温室气体排放量越多,生境适宜性下降的居群数目越多。
2.4 限制双花木属植物分布的主导气候变量
MaxEnt 3.2.19软件自动生成的各变量贡献率的结果(表1)表明,贡献率最大的前3个气候变量依次为最湿月降水量(bio13)、最干月降水量(bio14)和温度季节性变化标准差(bio4),分别为39.42%、25.69%、23.86%,累积值达88.97%。其中,与降水相关的最湿月降水量和最干月降水量的累积贡献率达65.11%。
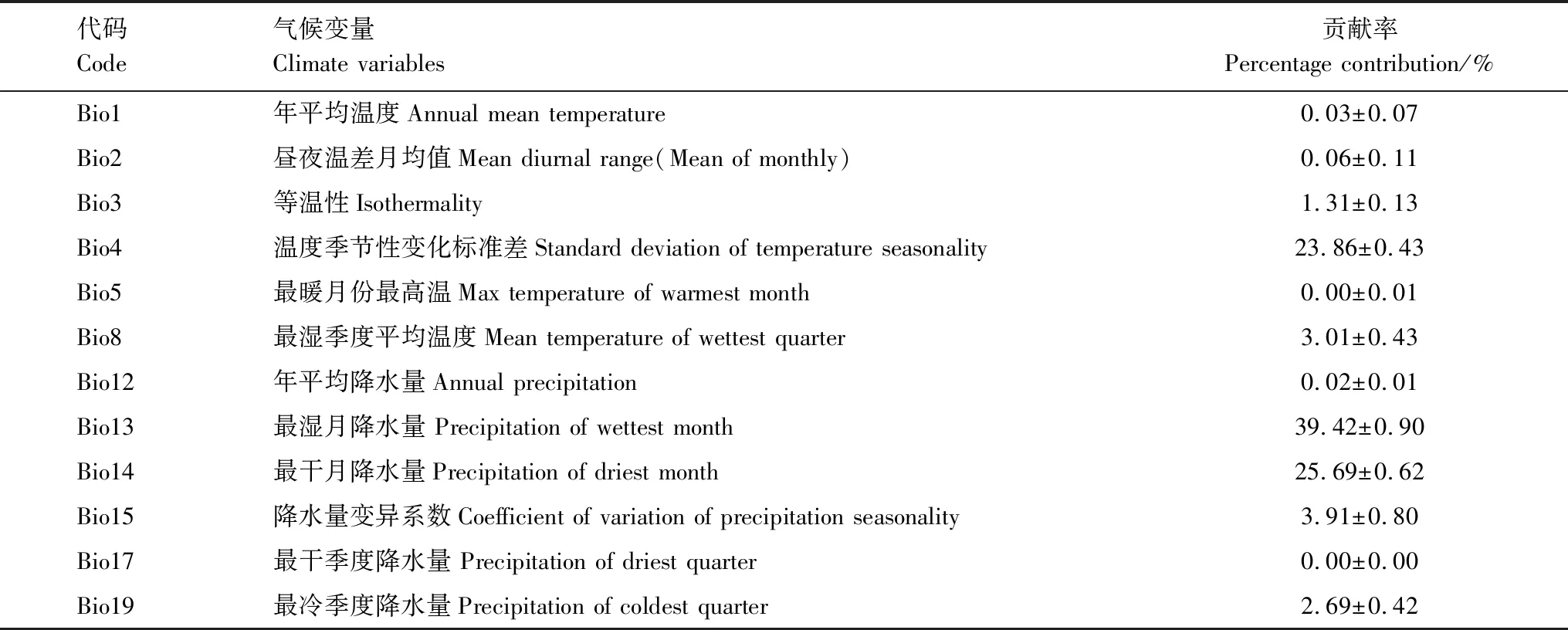
表1 用于模型预测的气候变量贡献率
刀切法检验结果(图2)显示,对双花木属植物分布影响较大的前5个气候变量:最冷季降水量(bio19)、最干月降水量(bio14)、最干季降水量(bio17)、年均降水量(bio12)及最湿月降水量(bio13)测试增益分别为2.05、1.91、1.86、1.78和1.61,相差不大。由此可见,限制双花木属分布的主导气候变量是与降水有关的最湿月降水量和最干月降水量。

图2 刀切法检验气候变量对双花木属分布影响的重要性Fig.2 The jackknife test on the contribution of climate variables to the distribution ofDisanthusMaxim.
2.5 气候变化对双花木属植物潜在分布的影响
MaxEnt 3.2.19软件的模拟结果表明,当前双花木属植物存在/不存在的阈值为0.39。当前与其他时期分布图叠加的结果(图3)显示,末次盛冰期以来(图3A),东海大陆架消失,中国的长柄双花木潜在分布区向东扩散,日本的双花木潜在分布区整体上收缩,但日本九州地区潜在分布区向北扩散。未来RCP 2.6和4.5气候情景下(图3B、3C),中国武夷山大部分地区、日本九州局部地区的气候条件不适宜双花木属植物生存;RCP 6.0(图3D)气候情景下,中国南岭山脉的长柄双花木适宜分布区面积大幅度减少,但日本的双花木适宜分布区面积基本不变;RCP 8.5(图3E)气候情景下,中国仅局部地区气候条件适宜长柄双花木分布,日本仅九州大部地区气候条件不适宜双花木分布。总体来说,未来温室气体排放量越多,双花木属植物潜在分布区收缩程度越大。
居群生境稳定性体现在气候变化对居群的影响,居群生境稳定性越高,气候变化对居群生存影响越小。由图3可知,末次盛冰期(图3A),中国莽山、蓝山、连州和日本冈山居群的生境稳定性高,中国道县、宜章、新宁、邵阳、宜丰和日本滋贺居群的生境稳定性中等,其他居群的生境稳定性低。未来,RCP 2.6气候情景下(图3B),中国邵阳及日本冈山、高知、滋贺、长野、歧阜居群的生境稳定性高。RCP 4.5气候情景下(图3C),中国邵阳、玉山、井冈山及日本冈山、高知居群的生境稳定性高。RCP 6.0气候情景下(图3D),中国龙泉、开化、玉山、井冈山、日本冈山、高知、广岛、滋贺、长野、歧阜居群的生境稳定性高。RCP 8.5气候情景下(3E),仅中国井冈山和日本歧阜居群的生境稳定性高。
采用ArcToolbox中的重分类工具,统计不同时期的双花木属植物适生分布区域的栅格数的结果见表2。

表2 不同时期双花木属植物分布区栅格数量比较
由表2可知,末次盛冰期双花木属Na比值为0.87,Ne为0.45;未来,在RCP 2.6、RCP 4.5、RCP 6.0、RCP 8.5的气候情景下,双花木属Na比值分别为1.32、1.23、1.42、2.40,Ne分别为0.34、0.30、0.37和0.65。有研究认为,Na接近1,意味着物种分布稳定;远低于1,潜在分布面积收缩;远高于1,潜在分布面积经历了扩张[26]。这表明,末次盛冰期以来,双花木属植物潜在分布区收缩,适生分布面积收缩了45%;与当前相比,未来,双花木属植物潜在分布区面积将发生不同程度(30%—65%)的收缩,其中,RCP 8.5气候情景下,双花木属潜在分布区面积收缩程度最大。
3 结论与讨论
3.1 MaxEnt模型模拟可靠性
本研究基于MaxEnt 3.2.19软件模拟的结果表明,ROC曲线下的面积AUC均值约为0.999,几乎接近于1,高于四照花Cornuskousasubsp.chinensis(0.8785)[28]、栓皮栎Quercusvariabilis(0.9175)[29]、山桐子Idesiapolycarpa(0.9560)[30]和节节麦Aegilopstauschii(0.9580)[31]。这表明,采用MaxEnt模型对双花木属植物潜在地理分布区模拟的效果好,可信度高。
有研究发现,随着样本量增大,MaxEnt模型预测精确度逐渐增加,最后趋于稳定,要使MaxEnt模型预测结果更加准确,样本量需根据不同的物种进行适当选取[32]。珍稀濒危特有物种的地理分布范围有限、生境要求特殊、居群数量小,自然历史信息有限,有可能导致模型不准确和过度拟合。为了准确预测珍稀濒危特有物种的适宜分布区,一般结合其近缘种居群数据进行模拟[33]。双花木属植物是生态位较窄的局域性物种,呈间断性分布。本研究综合使用长柄双花木与其原种双花木居群的数据进行模拟,预测的分布区与Flora of China(http://foc.eflora.cn/)及文献[17,19]记载基本一致。
采用ArcGIS 10.0生成的双花木属植物在不同时期的地理分布图(图1),直观展示了末次盛冰期、当前和未来不同气候情景下的潜在适宜分布区的变化。气候变量贡献率及刀切法结果显示,最干月降水量、最湿月降水量是双花木属植物分布的主导限制因子。这与野外实地调查的长柄双花木植物居群多数分布于沟谷、溪流两旁,温凉湿润环境的结果相吻合,说明环境湿度对双花木属植物的分布具有限制作用。
3.2 潜在分布区变化
在地理区域尺度范围,气候变量是决定物种分布的主要因素。气候变暖将引发冰川融化、海平面上升、区域降水改变和极端气候增多等现象,直接影响陆地植被分布的变化[34]。本研究结果显示,末次盛冰期以来,双花木属植物分布区面积收缩了45%,这与清香木(Pistaciaweinmannifolia)的研究结果相似(收缩44.4%)[35]。未来,随着温室气体的排放量增加,双花木属植物的潜在分布区将出现不同程度收缩,与乌冈栎(Quercusphillyraeoides)[36]的研究结果类似。本研究中,未来RCP 4.5气候情景下,双花木属的适宜分布区面积较RCP 2.6气候情景下的分布区面积略有增加,这可能与未来RCP 4.5气候情景下的温度季节性变异和极端低温事件大大减少有关[37]。未来双花木属适宜分布区面积减小,意味着未来气候变化下,双花木属植物的生存风险加剧,资源保护和管理面临严峻的挑战。
MaxEnt 3.2.19软件预测结果(图1B)显示,除了双花属植物现在已知居群外,中国黄山、武夷山山脉以及日本九州等地的生态气候也适宜双花木属生长分布。一种可能的解释是,这些地方极有可能存在双花木属植物分布,只是尚未被发现。事实上,中国湖南邵阳、蓝山、宜章和广东连州等地的长柄双花木居群是近年旅游开发过程中,开展植被本底调查时发现的。另一种可能的解释是,本研究仅基于物种居群的地理位置及气候因子,但物种分布除受气候因子影响外,还受物种间互作、迁移、地形、局部小气候等自然因子影响[38]。因此,要结合当地植被、土壤、地形地势、海拔,综合生物与非生物因子,才能有效地揭示物种真实的潜在分布区。
末次盛冰期,东海海平面下降120—130 m左右,绝大部分大陆架露出海面,中国与日本地区可能存在狭长连续森林的陆桥连接[39]。本研究结果显示,末次盛冰期陆桥的局部地区气候条件适合双花木属植物的生长。但是,基于cpDNA的双花木属系统发育的研究表明,中国的长柄双花木与日本的双花木无共享的单倍型[17],这可能是分布于陆桥的双花木属植物居群生境片段化,也可能是双花木属植物种子萌发受其自身因素限制和外界条件的干扰较大[40],阻碍了两大居群间的基因流。
3.3 资源保护
未来全球气候变暖,必然会改变双花木属植物的生长环境,影响其地理分布。研究濒危植物分布格局对气候变化的响应,可以掌握物种潜在生态适宜区的变化和面临的生态风险,对植物居群原地保护、人工引种、迁地保护地点的选择、野外资源调查点的指导及适宜分布区生境的保护和管理至关重要[11]。基于本研究结果(图1B),中国南岭、黄山、罗霄山、武夷山山脉及日本纪伊山、赤石山及越后山等地可确定为双花木属植物资源优先调查、保护地区,这将有助于发现双花木属新的居群以及提高迁地引种成活率。双花木属植物常分布于林缘的沟谷两边,由于经济发展,交通道路改善,原来人迹罕至的森林遭到更多的人为因素影响,双花木属植物居群破坏严重[18]。因此,对已发现的双花木属植物的现实居群,亟待在当地建立就地保护小区,通过定点观察和动态监测、分析,确定其居群和生境的变化规律,加强管理,确保其居群生存的安全。
目前,在中国井冈山和庐山两地已建立了长柄双花木迁地保护林[18],这对其物种保育具有重要的意义。但是,本研究模拟的结果表明,未来气候情景下(图3B—F),庐山气候条件不适宜长柄双花木生长、发育。建议加大专项经费投入,积极开展长柄双花木繁殖生物学、居群生态学和人工驯化等方面研究,促进庐山人工居群能够自然更新,避免遗传适应性变异、突变积累及遗传资源丧失等风险。
