外源氮持续输入对相应氮梯度下碱蓬残体分解及硫养分释放的影响
2019-05-31陈冰冰孙志高孙文广胡星云
陈冰冰,孙志高,*,孙文广,胡星云
1 福建师范大学湿润亚热带生态地理过程教育部重点实验室,福州 350007 2 福建师范大学地理研究所,福州 350007 3 福建师范大学福建省亚热带资源与环境重点实验室,福州 350007 4 路易斯安那州立大学植物、环境与土壤科学系, 巴吞鲁日 700803
河口湿地处于海陆相互作用地带,是海岸带地区响应全球变化和人类活动较为敏感的生态系统之一[1]。植物残体分解是河口湿地物质循环的重要环节,它连接着生物有机体的合成与分解[2],而分解过程中营养元素的动态变化直接影响着河口湿地的养分状况,进而影响到湿地系统的生产力。硫作为继氮、磷、钾之后的第四位营养元素是植物生长必需的中量矿质营养元素,在许多生物地球化学过程中扮演着重要角色,如参与蛋白质、氨基酸和叶绿素形成,控制光合作用中碳水化合物代谢、影响植物呼吸作用和抗逆性等[3],其对于维持河口湿地健康具有重要意义。目前,国内外关于河口湿地植物残体分解的研究大多集中在残体分解特征、影响因素(水分、盐度、温度、养分和沉积等)[4- 5]及元素释放(碳、氮、磷及痕量元素)[4,6- 7]等的探讨上,而关于养分条件改变对湿地植物残体分解及养分(特别是硫)释放的研究还不多见。
黄河口湿地是渤海与黄河河口相互作用形成的中国暖温带最广阔、最完整和最年轻的滨海湿地,承接着上游带来的大量含氮物质。碱蓬(Suaedasalsa)作为黄河口新生湿地的典型盐生植被,具有维持生物多样性、生态系统演替和调节气候等多重生态功能。近年来,黄河口的营养盐入海通量已达1.41—4.22×104t,并始终保持在较高水平[8]。另外,该区当前的氮湿沉降量为3—4.5 g m-2a-1[9],接近其氮湿沉降临界负荷(4.0 g m-2a-1)[10],而这均可能对湿地植物生长、残体分解及养分循环状况产生深刻影响[11]。当前,关于黄河口植物残体分解的相关研究主要集中于沉积强度及水盐梯度变化对残体分解与碳、氮及痕量元素释放的影响方面[6,12- 13],而关于外源氮输入对植物残体分解及养分释放的影响研究还不多见,特别是关于氮养分持续输入条件下湿地植物残体基质质量改变对残体分解速率和硫养分释放影响方面的研究还鲜有报道,而对此进行研究有利于预测未来黄河口氮养分增加条件下湿地氮、硫生物循环的耦合状况。鉴于此,本研究以黄河口北部滨岸高潮滩碱蓬残体为研究对象,将第一年度野外原位氮输入试验获得的不同基质质量的植物残体,分别原位投放至来源样区,并通过第二年度的持续输氮,探讨外源氮持续输入对相应氮梯度下碱蓬残体分解速率及硫养分释放的影响。研究结果有助于揭示黄河口潮滩碱蓬湿地氮、硫养分循环的耦合机制,并可为潮滩湿地的生态保育提供重要科学依据。
1 研究区域与研究方法
1.1 研究区概况
研究区位于山东省黄河三角洲国家级自然保护区(37°40′N—38°10′N,118°41′E—119°16′E),该保护区于1992年经国家林业局批准建立,是中国最大的河口三角洲自然保护区,在世界范围河口湿地生态系统中具有较强代表性,主要保护新生湿地生态系统和珍稀濒危鸟类。保护区总面积为15.3万hm2,属暖温带季风气候区,四季分明,雨热同期,冷热干湿界限明显,年降水量551.6 mm,70%的降水集中在7—8月,年均温12.1℃,无霜期196 d,≥10℃积温约4300℃,年蒸发量1928.2 mm。该区的土壤类型主要为隐域性潮土和盐土,主要植被类型为芦苇(Phragmitesaustralis)、碱蓬(S.salsa)、柽柳(Tamatixchinensis)、白茅(Imperatacylindrica)及罗布麻(Apocynumvenetum)等,其中芦苇、碱蓬和柽柳的分布较广。
1.2 研究方法
1.2.1试验设计
(1) 残体获取
2014年4—11月,选择今黄河入海口北部滨岸高潮滩的碱蓬湿地为研究对象,参照欧洲NITREX项目的研究设计,进行野外原位氮输入模拟试验。试验设4个处理,即N0[无额外氮处理,其值为当前湿地实际氮输入量(6.0 gN m-2a-1)。]、N1[低氮处理:1.5N0(9.0 gN m-2a-1),模拟未来较低的氮增加量]、N2[中氮处理:2.0N0(12.0 gN m-2a-1),模拟未来较高的氮增加量]和N3[高氮处理:3.0N0(18.0 gN m-2a-1),模拟未来更高的氮增加量],每个处理随机设置3个重复样地(5 m×10 m)。自4月下旬开始,每隔20—30 d以CO(NH2)2水溶液的形式对不同样地进行外源氮输入强度的模拟,共8次。对照样地喷洒等量的水以减少因外加水造成对湿地生态过程的影响。持续输氮一个生长季后,在植物生长季末(10月下旬),分别在上述N0、N1、N2和N3 4种氮输入样地内采集相应氮梯度下的植物残体(分别记为NL0、NL1、NL2和NL3)。
(2) 分解试验
将获取的植物残体带回实验室,剪成10 cm左右的小段,用蒸馏水清洗且烘干后,分别装进不同标记(用于区NL0、NL1、NL2和NL3)的分解袋中。分解袋由网眼为0.5 mm×0.5 mm的尼龙网制成,规格为20 cm×20 cm,每袋15 g(干质量)。四种氮输入处理下碱蓬残体的基质质量详见表1。
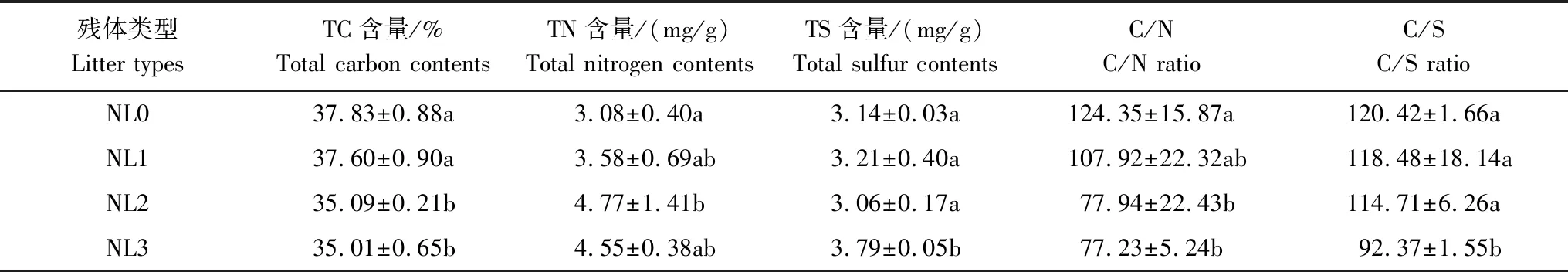
表1 不同基质质量碱蓬残体的基本性质
同列不同字母表示数据间在P<0.05水平上差异显著; NL0:无额外氮输入下的残体, Litters with no N import; NL1: 低氮输入下的残体, Litters with low N import; NL2: 中氮输入下的残体, Litters with middle N import; NL3: 高氮输入下的残体, Litters with high N import
2015年4月15日,将不同标记的残体分解袋投放至上述原位氮输入模拟试验的来源样区,即残体NL0投放至N0处理样区、NL1投放至N1处理样区、NL2投放至N2处理样区和NL3投放至N3处理样区,继续进行相应强度的输氮处理(方法与1.2.1.1原位氮输入模拟一致)。试验进行时,同步测定气温、地表温度、pH和电导率等环境因子(表2)。

表2 不同氮处理下碱蓬湿地环境因子
1.2.2样品采集与分析
试验期内(0—220 d),分别于2015年4月30日(第15 d)、5月25日(第40 d)、6月25日(第71 d)、7月25日(第101 d)、8月22日(第129 d)、9月24日(第162 d)、10月23日(第191 d)和11月21日(第220 d)进行了8次采样。将取回的分解袋及时带回实验室,去掉泥土并拣出袋内杂物,用去离子水将样品清洗干净后烘干至恒重。称量后,将样品用粉碎机磨碎,过0.25 mm筛,采用Vario EL 型元素分析仪测定样品的总碳(TC)、总氮(TN)和总硫(TS)含量。
1.2.3相关指标计算
残体残留率(R,%)和分解速率(d-1)可用下式计算:
ln(Wt/W0)=-kt
式中,W0(g)为残体干物质初始质量;Wt(g)为残体在t时刻的干物质质量。k为分解速率;t(d)为分解时间。
植物残体分解过程中硫的累积或释放可用硫累积指数(SAI)表示,即:
式中,DMi(g)为残体在i时刻的干物质质量;TSi(mg/g)为i时刻残体的TS含量;DM0(g)为残体干物质初始质量;TS0(mg/g)为残体初始TS含量。若SAI<100%,说明残体在分解过程中发生了硫净释放;若SAI>100%,说明发生了硫净累积。
1.2.4数据处理与分析
运用Origin 8.0软件对数据进行作图和计算。利用SPSS 23.0软件对不同氮输入处理下残体的残留率、TS和TC含量进行单因素方差分析,对残体的残留率与环境因子以及TS含量(或SAI)与残留率和环境因子进行Pearson相关性分析,对残体的残留率与环境因子以及TS含量与残留率、C/N、C/S和环境因子进行逐步线性回归分析。
2 结果与分析
2.1 氮输入对残体分解速率的影响

图1 不同氮处理下碱蓬残体残留率的变化 Fig.1 Variations of dry mass remaining ofSuaeda salsalitters in different nitrogen import treatmentsN0:无额外氮处理, No N import treatment; N1: 低氮处理, Low N import treatment; N2: 中氮处理, Middle N import treatment; N3: 高氮处理, High N import treatment
尽管不同氮输入条件下碱蓬残体的残留率在分解过程中均有所波动,但整体呈降低趋势(P>0.05)(图1)。分解220 d后,NL0、NL1、NL2和NL3在不同氮处理下的残留率分别为67.09%、56.24%、64.89%和57.73%。随着氮输入量的增加,不同基质质量残体的分解速率依次表现为NL1(0.002405 d-1)>NL3(0.002178 d-1)>NL2(0.001816 d-1)>NL0(0.001676 d-1)。与N0相比,N1、N2和N3处理下不同基质质量残体的分解速率分别增加了43.5%、8.35%和29.95%,而t0.95分别减少了1.43a、0.4a和1.09a(表3),表明外源氮持续输入条件下残体基质质量改变促进了残体的分解,且该促进作用在N1处理下表现的最为明显。
2.2 氮输入对残体中碳硫含量的影响
随着氮输入量的增加,尽管不同处理下残体分解过程中的TC含量整体均高于N0处理,但其在不同处理之间的差异均不显著(P>0.05)(图2)。0—101 d,残体中的TC含量整体呈上升趋势,尤以NL2和NL3在分解初期的增幅最为明显;101—162 d,NL0、NL2和NL3的TC含量均呈先增加后降低趋势,而NL1的TC含量呈一直降低变化;162—220 d,除NL0的TC含量呈持续增加外,其他残体的TC含量均呈先增加后降低变化。尽管不同氮处理下碱蓬残体在分解过程中的硫含量呈不同波动变化特征,但不同处理下残体TS含量之间的差异并不显著(P>0.05)(图2)。不同氮处理下残体的TS含量在0—70 d总体呈降低趋势,在70—162 d均呈较大波动变化,而在160—220 d均呈小幅增加趋势。220 d后,虽然NL0、NL1、NL2和NL3的TC和TS含量均相差不大,但其TC含量均高于相应处理的初始含量,而其TS含量则均低于相应处理的初始含量,降幅分别为22.61%、26.17%、28.34%和44.06%,说明随着氮输入量的增加,不同基质质量残体分解过程中的TC含量均呈增加趋势,而TS含量均呈降低趋势。
表3 不同氮处理下残体残留率的自然对数(y)与分解时间(t)的回归方程和参数
Table 3 Linear equations and parameters between the natural logarithm (y) of mass remaining and decomposition days (t) in different nitrogen import treatments

残体类型Litter types方程Equationsk (d -1)R2P(t0.95)/(a)NL0y=-0.08920-0.001676t0.0016760.82090.00084.68NL1y=-0.09352-0.002405t0.0024050.9296<0.00013.25NL2y=-0.10722-0.001816t0.0018160.73360.00324.28NL3y=-0.09250-0.002178t0.0021780.9232<0.00013.59

图2 不同氮处理下碱蓬残体全碳(TC)和全硫(TS)含量变化Fig.2 Variations of the total carbon(TC)and the total sulfur(TS)contents inSuaeda salsalitters in different nitrogen import treatments
2.3 氮输入对碳硫累积与释放的影响
不同氮处理下不同基质质量残体的碳和硫养分在分解期间均表现为不同程度的净释放(图3)。其中,碳释放量(CAI)随分解进行整体呈增加趋势,且以N1处理最高,N0处理次之,N2和N3处理较低。硫释放量随分解进行整体呈波动增加变化,其SAI在N0、N1、N2和N3处理下分别降低了15.26%—59.04%、15.11%—69.69%、15.80%—64.47%和27.25%—73.78%。0—70 d,N3处理下残体的硫养分释放最多,且释放量远高于其他氮处理;70—162 d,N1处理下残体的硫养分释放最多,而N2和N3处理下的硫养分释放较少;162—220 d,除N1处理外,其他处理下残体中的硫养分均呈增加趋势。220 d后,不同基质质量残体的碳释放量表现为NL1>NL3>NL0>NL2,而硫释放量表现为NL3>NL1>NL2>NL0,说明外源氮持续输入条件下残体基质质量改变对碳硫养分释放均具有一定的促进作用,且该作用分别在N1和N3处理下表现的最为明显。

图3 不同氮输入处理下碱蓬残体碳硫累积指数(CAI、SAI)的变化Fig.3 Variations of carbon and sulfur accumulation indices(CAI and SAI)ofSuaeda salsalitters in different nitrogen import treatments
2.4 不同氮处理下主要参数相关分析
不同氮处理下残留率与环境因子的相关分析表明,N2和N3处理下持续输氮导致的残留率与pH呈显著负相关(P<0.05),N0、N2和N3处理下的残留率与EC呈显著或极显著正相关(P<0.01或P<0.05),而其他环境因子(湿度、气温、地表温度)与不同处理下残留率之间的相关性均未达到显著水平(P>0.05)(表4)。另外,不同氮处理下残体的碳硫含量及累积指数(TC、TS、CAI、SAI)与残留率、C/N比、C/S比和不同环境因子的相关分析表明,除N1和N2处理下残体的TC含量与pH呈显著负相关(P<0.05),与气温呈显著正相关外(P<0.05),其他相关因子与其的相关性均未达到显著水平(P>0.05)(表5)。不同氮处理下残体的TS含量与C/S比之间均呈极显著负相关(P<0.01),仅N2处理下的TS含量与残留率和EC呈显著正相关(P<0.05)。相比之下,4种氮处理下的CAI大多均与残留率和C/N比呈极显著或显著正相关(P<0.05或P<0.01),N1和N3处理下CAI与EC呈显著正相关(P<0.05),N2和N3处理下CAI与pH呈显著负相关(P<0.05)。不同氮处理下残留率、C/N比和C/S比是影响残体SAI的主要因素,其中残留率与SAI均呈显著或极显著正相关(P<0.05或P<0.01);除N3外,其他氮处理下C/N比与SAI均呈显著正相关(P<0.05),而C/S比则与其呈极显著负相关(P<0.01)。此外,N2和N3处理下pH与SAI呈显著负相关(P<0.05),而N2处理下EC与SAI呈极显著正相关(P<0.01)。

表4 不同氮处理下残体残留率与环境因子之间的相关性
*为P<0.05;**为P<0.01

表5 不同氮处理下碳硫参数(TC、TS、CAI、SAI)与各因子之间的相关性
*为P<0.05;**为P<0.01
3 讨论
3.1 氮输入对残体分解的影响
本研究表明,不同氮处理下碱蓬残体的残留率之间不存在显著差异(P>0.05),这可能与不同氮输入研究样地的环境因子之间差异不大有关(表2)。前述分析表明,除N2和N3处理下残留率与pH呈显著负相关以及N0、N2和N3处理下残留率与EC呈显著或极显著正相关外,不同处理下残留率与其他环境因子之间的相关性均不显著(表4)。逐步线性回归分析进一步表明,除N1外,EC(x)均是导致其他氮处理下残留率变化的主要影响因素(N0:y=64.374+0.342x,R2=0.526,P=0.04;N2:y=59.072+0.413x,R2=0.684,P=0.011;N3:y=54.265+0.487x,R2=0.768,P=0.004)。另据图4可知,不同研究样地,持续氮输入导致表层土壤的pH和EC均发生了较为明显的变化,特别是持续输氮100 d后,相对于N0处理,pH值均随氮输入量的增加而降低,而EC值则随其输入量的增加而升高。已有研究表明,pH过高或过低均会对微生物活性产生抑制作用,进而不利于枯落物分解的进行[14]。James和Robert的研究还表明,增加酸度可抑制Sparganiumeurycarpum枯落物的分解,且残留率随着pH的降低而相对增加[15],说明枯落物的分解过程受到抑制。本研究结果与之不同,即随着氮输入量的增加,尽管pH呈降低趋势(图4),但残体分解速率整体呈增加趋势(表3),说明其他因素在残体分解中起到了促进作用,而这一因素可能与EC密切相关。一般而言,盐度(以EC表征)增加可抑制残体分解[5,16],但本研究却得到相反的研究结果,即随着氮输入量的增加,EC呈增加趋势(图4),而残体分解速率整体随之加快(表3)。Mendelssohn等在日德兰半岛北部自然盐分梯度带上对纤维素分解程度的研究也得到类似结论,即在最高盐分处,每日的CTSL(Cotton Tensile Strength Loss)显著增加[17]。Sun等在研究黄河口潮滩碱蓬残体分解对不同沉积强度响应时也得到类似的结果,并指出高盐分(以EC表征)条件下,残体较高的分解速率可能与适应于高盐分微生物的活性有关[18]。Lopes等研究进一步发现,在盐度较高的潮间带,黑角藻(Fucusvesiculosus)的分解速率一般较高,而这的确与一些耐高盐分的微生物在盐分较高条件下的活性较强有关[5]。

图4 不同氮输入处理下土壤表层pH和电导率(EC)的变化Fig.4 Variations of pH and electric conductivity(EC)of surface soils in different nitrogen import treatments

图5 不同氮输入处理下碱蓬残体C/N比和C/S比的变化 Fig.5 Variations of C/N and C/S ratios inSuaeda salsalitters in different nitrogen import treatments
不同氮处理下碱蓬残体分解速率的变化不仅与持续输氮导致的上述pH和EC的变化有关,而且还可能与残体本身的初始基质质量以及持续输氮条件下残体基质质量的变化有关。已有研究表明,残体自身的基质质量是影响其分解的主要因素。一般而言,生长在富养分环境中的植物,其残体的C/N比越低,耐分解化合物的含量相对就较少,分解越快,反之越慢[19]。本研究主要是基于第一年度野外原位输氮试验获得的植物残体,通过原位投放至相应来源样区,探讨第二年度持续氮输入条件下碱蓬残体分解速率的差异。本研究表明,不同氮输入条件下碱蓬残体的分解速率均高于N0处理,即氮输入梯度明显促进了残体的分解。由于第一年度外源氮输入的持续增加,改变了植物体内的C、N含量及其他元素含量,从而使得不同氮处理下植物残体的基质质量在本研究初期就产生了明显分异,且随着氮输入量的增加,C/N比和C/S比均呈降低趋势(表1)。另外,NL2和NL3的C/N比与NL0的C/N比,以及NL3的C/S比与 NL0、NL1和NL2的C/S比均存在显著差异(P<0.05)。与NL0相比,NL1、NL2和NL3处理下残体在分解初期的C/N比均明显低于NL0,由此可在很大程度上解释不同氮处理下残体分解速率均高于N0处理的原因。相关研究也得到类似的结果。Verhoeven和Toth的研究表明,富养分沼泽中生长的圆锥苔草(Carexdiandra)的枯落物比贫养分沼泽中生长的丘间种喙叶泥炭藓(Sphagnumfallax)枯落物的分解快[20]。Aerts和Caluwe的研究发现,增加氮沉降可促进苔草(Carex)枯落物的分解[21]。Song等研究还发现,氮输入增加了小叶章(Calamagrostisangustifolia)残体的氮含量,降低了其C/N比,促进了其残体分解[22]。本研究还表明,第二年度持续氮输入条件下具有不同基质质量碱蓬残体的分解速率整体表现为NL1>NL3>NL2>NL0,即相对于N0处理,N1处理最高,N3处理次之,N2处理最低,原因可能与不同氮输入梯度下残体分解过程中C/N比改变程度的差异有关。此外,外源氮输入在改变残体基质质量的同时,也可提高了微生物的活性,特别是在高C/N比时,微生物可通过吸收大量外源氮来满足自身生长的需要,进而加快残体的分解[22]。Vivanco和Austin等的研究也发现,氮养分供给充足能够刺激微生物的活性,从而加速残体的分解[23]。本研究中,尽管外源氮输入在分解过程中能够为微生物提供氮源而促进分解,但随着氮输入量的持续增加,氮养分对残体分解的影响程度可能在降低。由于氮是黄河口潮滩碱蓬湿地的重要限制性养分[18],且NL1的初始C/N比与 NL0的初始C/N比相差不大(P>0.05)(表1),而NL1的C/N比变化在第二年度对养分改变(低氮处理)的响应程度更为明显(图5),由此导致其分解过程中微生物的活性可能增强,分解速率较高。与之相比,尽管NL2和NL3的初始C/N比远低于NL0,但随着分解过程的进行, NL2和NL3的TC含量在分解后期明显高于NL0(图2),这就使得其C/N比在分解后期整体呈增加变化(图5),而C/N比的增加可对残体分解产生一定的抑制作用[19]。另外,相对于N1处理,尽管N2和N3处理下具有较强的氮供应强度,但其相应残体在分解后期由于C/N比的增加微生物对氮的需求量可能并不高[24],由此导致两种氮处理下残体(NL2和NL3)的分解速率低于NL1。与NL2相比,尽管NL3的分解速率随着氮输入量的增加呈一定的加快趋势,但相对于NL1仍较低,说明随着氮输入量的增加,碱蓬残体分解对养分供给的响应程度表现为不敏感或影响不大。James和Robert的研究也得到类似的结果,即增加氮供给对Sparganiumeurycarpum枯落物分解并无明显影响[15]。Xie 等对凤眼莲(Eichhorniacrassipes)、苦草(Vallisnerianatans)和黄丝草(Potamogetonmaackianus)三种水生植物叶片分解对氮供给响应的研究也发现,增加氮可利用性对三者分解的影响均不大[25]。
3.2 氮输入对残体碳硫含量及累积与释放的影响
前述分析可知,除N1处理下的TC含量与pH呈显著负相关以及N2处理下的TC含量与气温(T)呈显著正相关外,其他因子与TC含量的相关性均未达到显著水平(表5);而逐步线性回归分析也得到类似结果(NL1:y=-62.289pH+892.745,R2=0.597,P=0.025;NL2:y=2.185T+352.203,R2=0.654,P=0.015),说明不同氮处理下影响残体TC含量变化的因素可能较为复杂。Sun等对黄河口新生湿地芦苇、碱蓬(S.salsa)与盐地碱蓬(S.glauca)残体分解的相关研究也得到类似结论[6]。本研究还表明,不同氮处理下残体的CAI在分解期间均一直呈释放状态。相关分析发现,4种氮处理下残体的CAI大多均与残留率和C/N比呈极显著或显著性正相关(表5),说明残留率与C/N比是影响残体CAI变化的重要因素。比较而言,不同氮处理下残体的CAI整体以NL1最高,NL0次之,NL2和NL3相对较低,说明导致残体CAI变化的因素除残留率、C/N比外,还可能取决于其自身分解速率以及不同氮处理下其他环境因子的变化。如NL1残体的CAI最高,可能与该处理下CAI与EC呈显著正相关(表5)且其分解速率最快有关(表3);NL2和NL3残体的CAI均较低,可能在于两种处理下CAI与pH均呈显著负相关(表5),而这正好与前述持续氮输入条件下EC增加和pH降低促进了残体分解的研究结果相对应。
本研究发现,不同氮处理下碱蓬残体中TS含量均呈不同程度的波动变化特征,且TS含量在分解过程中均出现阶段性升高(图2),这可能与微生物对分解环境中硫的固持作用有关[26]。Sun等对无外源氮输入条件下黄河口潮滩湿地碱蓬残体分解的研究表明,残体分解过程中硫含量的阶段性升高主要与微生物对海水和沉积物中硫的固持作用有关[27]。李新华等在研究三江平原小叶章枯落物分解时亦表明,枯落物中硫含量的明显增加主要源于微生物对沼泽水中硫的固持作用[28]。尽管不同氮处理下残体TS含量的变化并不一致,但其均与相应的C/S比呈相反变化趋势。前述分析表明,不同氮处理下残体中TS含量与C/S比之间均呈极显著负相关(表5),说明C/S比可能是调控不同氮输入处理下残体分解过程中硫含量变化的主控因素。逐步线性回归分析结果还表明,C/S比(x1)是影响NL0分解过程中硫含量(y)变化的主控因素(y=5.055-0.014x1,R2=0.732,P=0.007),其对硫含量变化的解释贡献率高达73.2%。与之不同,C/S比和残留率(x2)共同影响着NL1分解过程中硫含量(y)的变化(y=2.712-0.009x1+0.019x2,R2=0.971,P=0.001),C/S比、气温(x3)和C/N(x4)比是影响NL2分解过程中硫含量(y)变化的三个重要因素(y=4.134-0.013x1+0.017x3+0.005x4,R2=0.997,P=0.000),而C/S比、C/N比和EC(x5)在影响NL3分解过程中硫含量(y)变化中的贡献最大(y=5.541-0.012x1-0.026x4+0.005x5,R2=0.996,P=0.000)。上述结果进一步说明了C/S比对不同氮处理下残体分解过程中硫含量变化的重要影响。Sun等以及李新华等在研究无氮输入条件下黄河口潮滩碱蓬残体以及三江平原小叶章残体分解过程中硫含量变化时亦得到类似结论[27-28]。
本研究还发现,持续氮输入条件下不同基质质量残体在分解过程中均表现为不同程度的硫释放,且其释放特征与相应残留率的变化趋势较为一致(图1和图3)。前述分析还表明,不同基质质量残体的残留率与相应的SAI均呈显著或极显著正相关(表5),说明不同氮处理下残体分解过程中的质量损失对其硫养分释放具有重要影响。另外,C/S比和C/N比也是影响残体SAI变化的两个重要因素。由于不同氮处理下残体的C/S比与TS含量密切相关(表5),故其对相应残体的SAI亦可产生重要影响。值得注意的是,除NL3外,NL0、NL1和NL2的C/N比与SAI均呈显著正相关(表5),原因可能是与分解期间微生物对持续高氮输入响应不敏感,使得C/N比对残体SAI的影响减弱有关。整体而言,分解前期(0—40 d),NL3的SAI明显高于其他残体。由于此间不同氮处理下残留率变化比较一致且其值也较为接近,故NL3在此间的较高硫养分释放强度主要取决于其较高的硫初始含量(表1)。分解中期(40—162 d),NL1的SAI最高,原因主要与其在持续氮输入条件下的分解速率最高有关。分解后期(162—220 d),不同基质质量残体的SAI表现为NL3>NL1>NL2>NL0,说明不同强度的外源氮持续输入均促进了相应残体的硫养分释放。由于NL2的分解速率在3种氮处理中最低,故其SAI相对也较低。尽管NL1的分解速率在3种氮处理中最快,但其SAI却低于NL3,原因可能在于NL1的硫养分释放量主要取决于其干物质损失量,而NL3的硫养分释放量除了取决于残体干物质损失量外,还可能与分解过程中微生物对硫的固持作用减弱有关。虽然黄河口潮滩碱蓬湿地主要受氮养分限制[18],但随着氮输入量的持续增加,氮不再是影响碱蓬残体分解过程中的主要限制性元素,特别是在高氮输入条件下(N3处理),微生物对N、S等的需求量也会降低[29],由此导致分解后期残体中多余的硫养分被释放出来,从而导致NL3的硫养分释放量较高。
近年来,黄河口的营养盐入海通量尽管年际变化较大,但一直保持在较高水平[8],这就使得黄河口潮滩湿地的氮养分状况可能会不断被改善。在未来黄河口氮负荷增强的情况下,碱蓬残体的基质质量将会被改变,而氮负荷的持续增加又会促进不同基质质量残体中硫养分的归还。因此,从硫养分归还角度可知,黄河口潮滩湿地氮负荷量的增加在一定程度上加速了其硫生物循环速率,其对于增加植被的氮硫养分供给以及维持潮滩湿地的稳定与健康具有重要意义。
4 结论
(1)不同基质质量残体的分解速率整体表现为NL1>NL3>NL2>NL0,说明外源氮持续输入条件下基质质量改变促进了残体的分解,且该促进作用在低氮处理下表现的最为明显,原因主要与其在残体分解过程中C/N比的改变程度最为明显有关。
(2)不同基质质量残体中的TS含量均呈不同程度的波动变化特征,且其变化特征均与相应的C/S比呈相反变化趋势,说明C/S比可能是调控不同氮持续输入条件下残体分解过程中硫含量变化的主控因素。
(3)随着氮输入量的增加,不同基质质量残体的硫养分在分解期间均发生不同程度的净释放,且释放强度整体表现为NL3>NL1>NL2>NL0,说明外源氮持续输入条件下残体基质质量改变促进了其硫养分的释放,且该促进作用在高氮处理下表现的最为明显。
(4)未来黄河口氮养分负荷增加的情况下,碱蓬残体的C/N比和C/S比将发生改变,而持续增强的氮负荷又会促进不同基质质量残体的硫养分归还,从而加速硫生物循环速率。
