乌梁素海冻融前后浮游植物群落结构特征及其影响因素分析
2019-05-31孙鑫李兴勾芒芒
孙鑫,李兴 ,勾芒芒
1. 内蒙古师范大学地理科学学院,内蒙古 呼和浩特 010022;2. 内蒙古大学生态与环境学院,内蒙古 呼和浩特 010021;3. 内蒙古师范大学节水农业工程研究中心,内蒙古 呼和浩特 010022;4.内蒙古机电职业技术学院水利与土木建筑工程系,内蒙古 呼和浩特 010070
浮游植物作为水生态系统的初级生产者以及水生态系统物质循环和能量流动的基础,能够对水体的状态变化做出迅速响应(韩博平等,2003)。目前,学者大多对中国东南部平原湖区和西南部云贵高原湖区进行浮游植物群落及其影响因素的研究,这些湖区温度最低也在0 ℃以上,不存在冻融情况(谢斌等,2017;Shi et al.,2016),而对存在冻融情况的北方寒旱区湖泊水体和冰体中浮游植物群落与营养盐分布的研究尚不多见(李卫平等,2014)。中国北方湖区在每年 11月份,水面开始结冰,非冰封期转为冰封期(李畅游等,2014)。水体表面冻结成冰后,水体的营养盐分布和水动力情况等发生改变,故水体经冻结后营养盐的迁移分布、浮游植物群落特征也会发生变化(李兴等,2018)。对北方寒旱区湖泊开展冻融前后的浮游植物群落特征变化研究十分必要,一方面全面研究代表蒙新高原湖区的乌梁素海对保护寒区湖泊环境具有必要性,另一方面拓宽在冰体环境研究方面的内容,对保护西北生态环境具有十分重要的意义。
乌梁素海位于内蒙古自治区乌拉特前旗,地理坐标为 40°36′-41°03′N,108°43′-108°57′E,为内蒙古西部最大的淡水湖泊。乌梁素海地处蒙新高原,每年承接河套平原地区大量农田退水,生态功能十分重要,为黄河中上游重要的保水、蓄水和调水场所(李兴等,2015)。2002年被国际湿地公约组织正式列入《国际重要湿地名录》(孙鑫鑫等,2009)。乌梁素海每年11月冰冻,翌年3月融化,具有较长的冰封期(吕宏洲等,2015),特殊的地理位置与气候环境使得乌梁素海浮游植物群落特征及其影响因素与其他区域湖泊有所不同。因此,对 2016年 10月、11月以及2017年1月、4月和5月乌梁素海湿地冻融前后浮游植物群落特征及其影响因素展开研究,为寒区湖泊浮游植物结构特征变化机理研究以及寒区湖泊环境保护提供科学依据。
1 材料与方法
1.1 采样点布置
根据乌梁素海水生植物、水动力特征及入出湖口等情况,在空间上将乌梁素海以2 km×2 km的正方形网格进行剖分,在网格的交点处以梅花形布设取样监测点12处(图1),其中小海子区域人为干预少、水深小于0.5 m,且芦苇密集,采样船无法到达,故不设监测点。采样点包括进水口J11、I12,出水区域河口HK,北部明水区域L15,芦苇区域N13,旅游区域 Q10、Q8,西大滩 L11,大卜洞 O10,大北口DBK,海壕HH,二点ED。
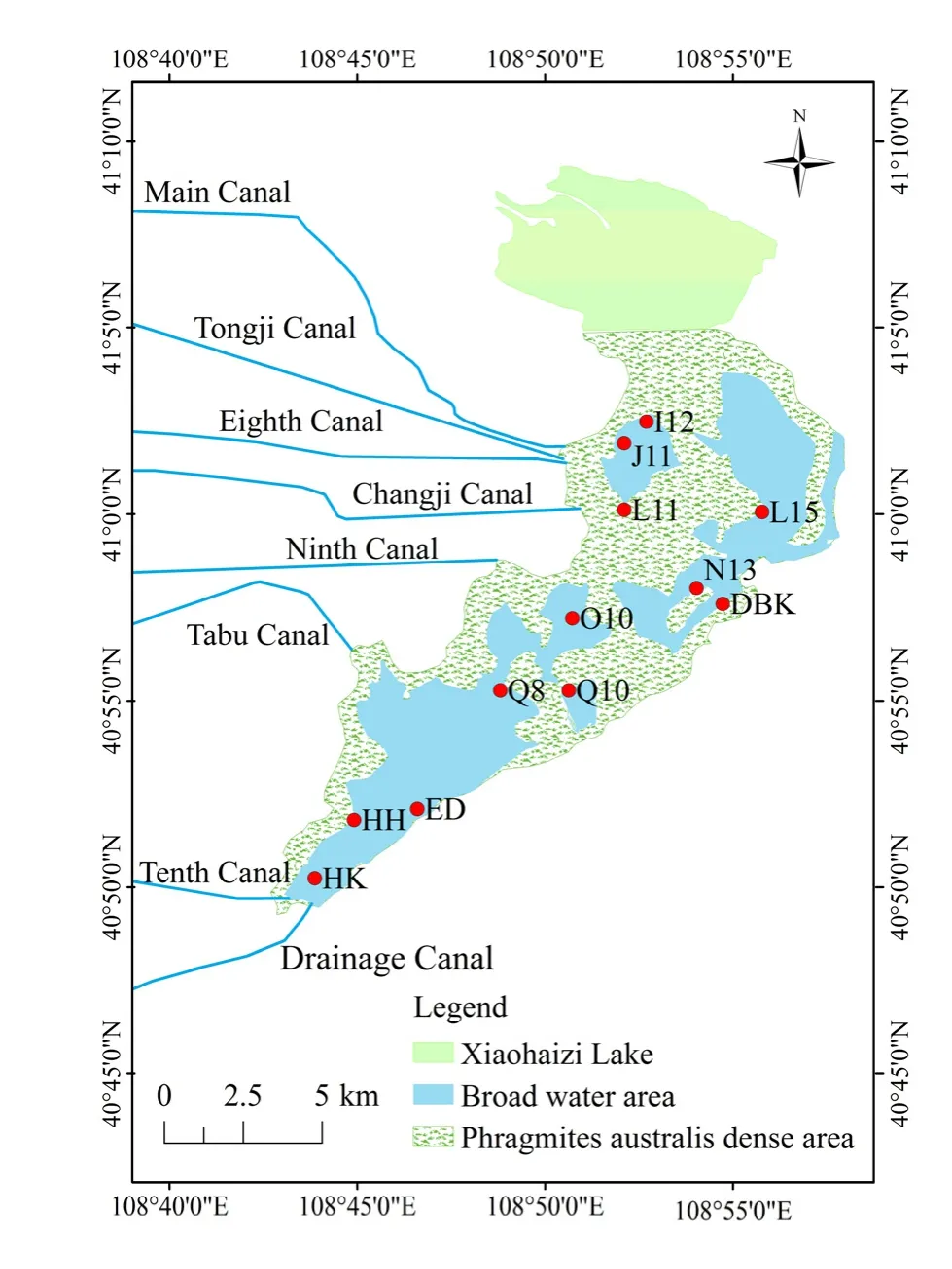
图1 乌梁素海浮游植物采样点分布示意图Fig. 1 Schematic diagram of monitoring site of phytoplankton in Wuliangsuhai
1.2 样品采集与数据处理
本研究采样时间为2016年10月、11月和2017年1月、4月、5月。因乌梁素海平均水深小于3 m,故在表层以下0.5 m处采样即可。参照《湖泊生态调查观测与分析》(黄祥飞等,1999),现场测定水深(WD)、水温(t)、pH、透明度(SD)、电导率(EC)、溶解氧(DO)、氧化还原电位(ORP)、盐度(SAT)、总溶解性固体(TDS)等参数;总氮(TN),总磷(TP)、氨氮(NH4+-N)、硝酸氮(NO3--N)、亚硝酸氮(NO2--N)、化学需氧量(COD)、叶绿素(Chl.a)、悬浮物(SS)等其他水质指标需采集1 L水样带回实验室进行测定,测定方法参照《水和废水监测分析方法》(第4版)(国家环境保护总局水和废水监测分析方法编委会,2002)。于2017年1月(冰封期)冰层稳定期采集冰样,用冰钻采集器破冰后,因不同采样点结冰厚度存在差异,为了确保数据的准确性、完整性,各个采样点酌情将冰层按每层10 cm分3-5层,分别置于2000 mL塑料瓶内,及时送回实验室,在室温条件下自然融化后取1000 mL水样进行水质测定以及浮游植物的镜检分类,同时用冰钻采集器破冰后采集 1 L冰下水样带回实验室也进行水质测定及浮游植物镜检分类。
浮游植物定性样品使用 25号浮游生物网按照“∞”形捞取,用4%甲醛溶液固定后带回实验室用于镜检分类。定量样品用采水器采集1 L水样后,加鲁哥试剂固定,将采集的浮游植物样品带回实验室后静止、沉降、浓缩至30 mL,摇匀取0.1 mL浓缩样品置于0.1 mL计数框内,在400倍显微镜下镜检,鉴定浮游植物种类及计算细胞丰度,参照《中国淡水藻类——系统、分类及生态》(胡鸿钧等,2006)及《淡水微型生物图谱》(周凤霞等,2005)等进行浮游植物种类的鉴定。
采用Margalef丰富度指数(D)、Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J)以及优势度(Y)指数(Belaoussoff et al.,2005;Lampitt et al.,1993)评价研究区域浮游植物多样性,计算公式如下:
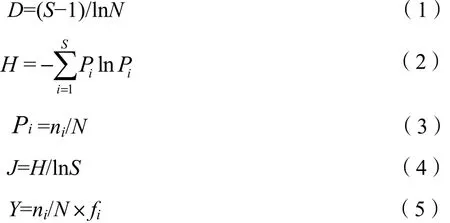
式中,Pi为第i种藻类的个数与样品中所有藻类个数的比值;ni为第i种藻类的个数;N为所有藻类总个数;S为样品中藻类种类数;fi为第i种藻类在各站位出现的频率。本文将优势度Y>0.02的藻类定为优势种。
采用ArcGIS 10.2统计模块作为空间分析工具,以反距离插值方法进行区域插值,对浮游植物丰度进行空间分布趋势模拟。采用CANOCO 4.5软件对物种数据和环境数据进行冗余分析,其中物种数据采用浮游植物丰度指标,按照物种至少在一个样点出现的频度>12.5%且至少在一个样点的相对丰度≥1%进行筛选,以降低稀有物种权重,物种矩阵与环境数据(除pH)进行lg(x+1)转换(Lopes et al.,2005;Muylaert et al.,2000)。
2 结果
2.1 种类组成
2016年10月、11月以及2017年1月、4月和5月共鉴定出浮游植物69属120种,隶属于蓝藻门(Cyanophyta)、绿藻门(Chlorophyta)、硅藻门(Bacillariophyta)、隐藻门(Cryptophyta)、裸藻门(Euglennophyta)和甲藻门(Pyrrophyta)。浮游植物群落以硅藻门最多,为 34属 50种,占总种类的41.67%,其次为绿藻门,为23属31种,占总种类的25.83%,蓝藻18属29种,占总种类的24.17%,裸藻3属7种,占总种类的5.83%,隐藻门2属2种,甲藻门1属1种,共占总种类的2.50%(图2)。种类数以冰封期1月份最多,为117种,10月、5月次之,分别为87、83种,11月与4月的种类数相差不大,分别为76和74种。分析浮游植物6门种类数随月份变化可知(图3),乌梁素海浮游植物冻融前后硅藻种类数均占主导地位,且与总种类数随月份的变化趋势相似。硅藻能适应低温环境条件,调查期间所有的硅藻种类均在冰封期1月份出现,且冰层硅藻种类大于水层硅藻种类;蓝藻与绿藻种类数次之,随月份的变化较小;金藻、裸藻、隐藻以及甲藻种类数所占比例很小,月份间变化不大。
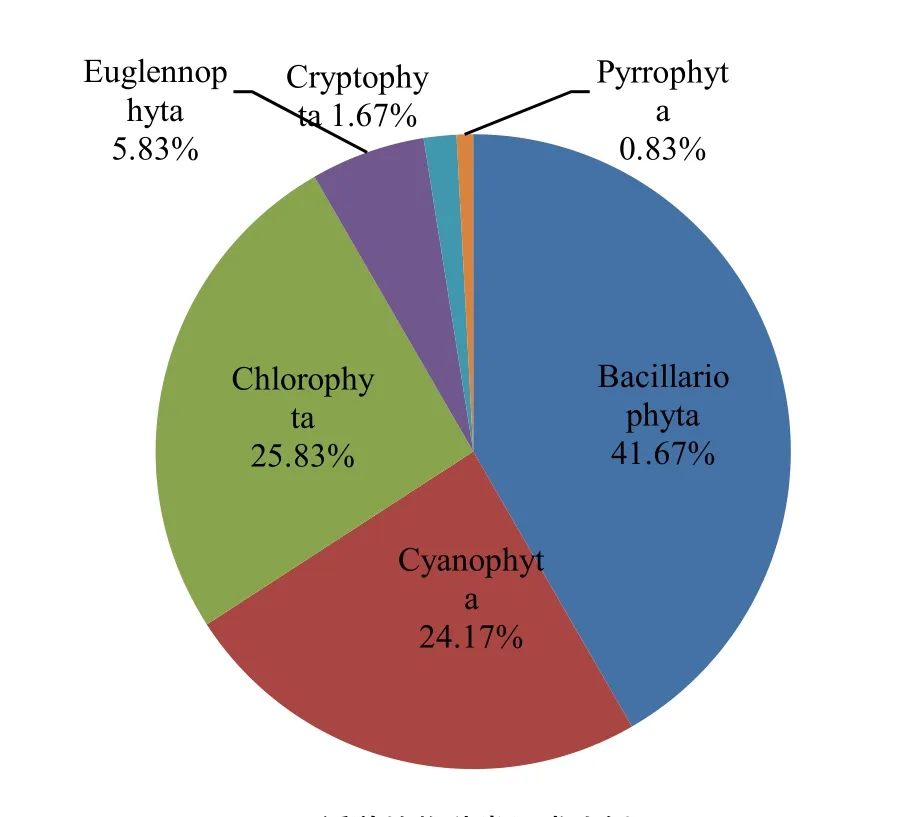
图2 浮游植物种类组成比例Fig. 2 Composition ratio of phytoplankton species
2.2 优势种
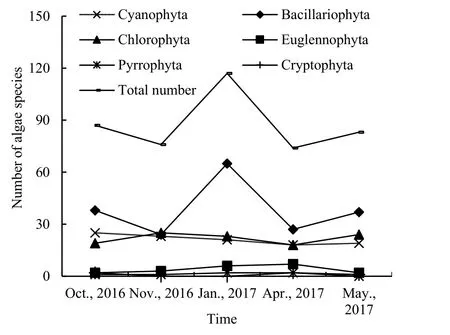
图3 乌梁素海浮游植物各门种数量随时间变化过程Fig. 3 Variation of the number of phytoplankton species in Wuliangsuhai over time
乌梁素海冻融前后浮游植物优势种、优势度见表1。由表1可知,冻融前后浮游植物优势类群差异较大。10月硅藻门、蓝藻门优势度较大,其中蓝藻门的优势种类最多,以蓝藻门的微小平裂藻(Merismopedia tenuissima)的优势度最大(0.173),11月硅藻门、蓝藻门、绿藻门优势度较大,其中以硅藻门的梅尼小环藻(Cyclotella meneghiniana)(0.120)的优势度最大,冰封期1月硅藻门优势种占绝对优势,以硅藻门丰度最高,其中硅藻门的双头辐节藻(Stauroneis anceps)(冰层:0.220)和尖针杆藻(Synedra acus)(冰层:0.132;水层:0.180)的优势度最大,结冰消融后的 4、5月则以绿藻、蓝藻为优势门类,蓝藻门的微小平裂藻(Merismopedia tenuissima)(4 月:0.073;5 月:0.155)和绿藻门的双对栅藻(Scenedesmus bijuga)(4月:0.089;5月:0.121)的优势度最大。
2.3 浮游植物丰度的时空变化
乌梁素海2016年10、11月和2017年1、4、5月各月平均丰度分别为 2.90×107、4.31×106、1.46×107、1.14×107、4.50×107cells·L-1。由图 4 可知,10月浮游植物丰度较大,11月丰度最小,次年逐渐增多,冰封期1月水层浮游植物丰度为冰层浮游植物丰度的1.30倍,分别为8.26×106、6.36×106cells·L-1,春季5月丰度最大。不同门类浮游植物丰度随月份的变化具有显著规律性,乌梁素海冻融前后的几个月浮游植物丰度均以蓝藻、绿藻以及硅藻为主,但各月随温度变化所占比例差异明显。10月所占比例最大为蓝藻(78.19%),其次为绿藻(12.68%);11月蓝藻(55.23%)比例最大,但相比10月有所下降,其次为硅藻(23.55%),硅藻相比10月丰度剧增;次年冰封期1月硅藻(63.02%)比例最大,其次为绿藻(16.16%)、蓝藻(13.27%);4月硅藻(40.62%)比例最大,相比1月硅藻比例有所下降,蓝藻与绿藻有所上升;5月蓝藻比例最大(64.52%),相比4月蓝藻急剧增多,硅藻减少。浮游植物种群结构在冻融前以蓝藻占优势,随着冰封期的到来,蓝藻占比渐少,硅藻占比渐多,冰封期后蓝藻占比渐多,硅藻占比渐少。
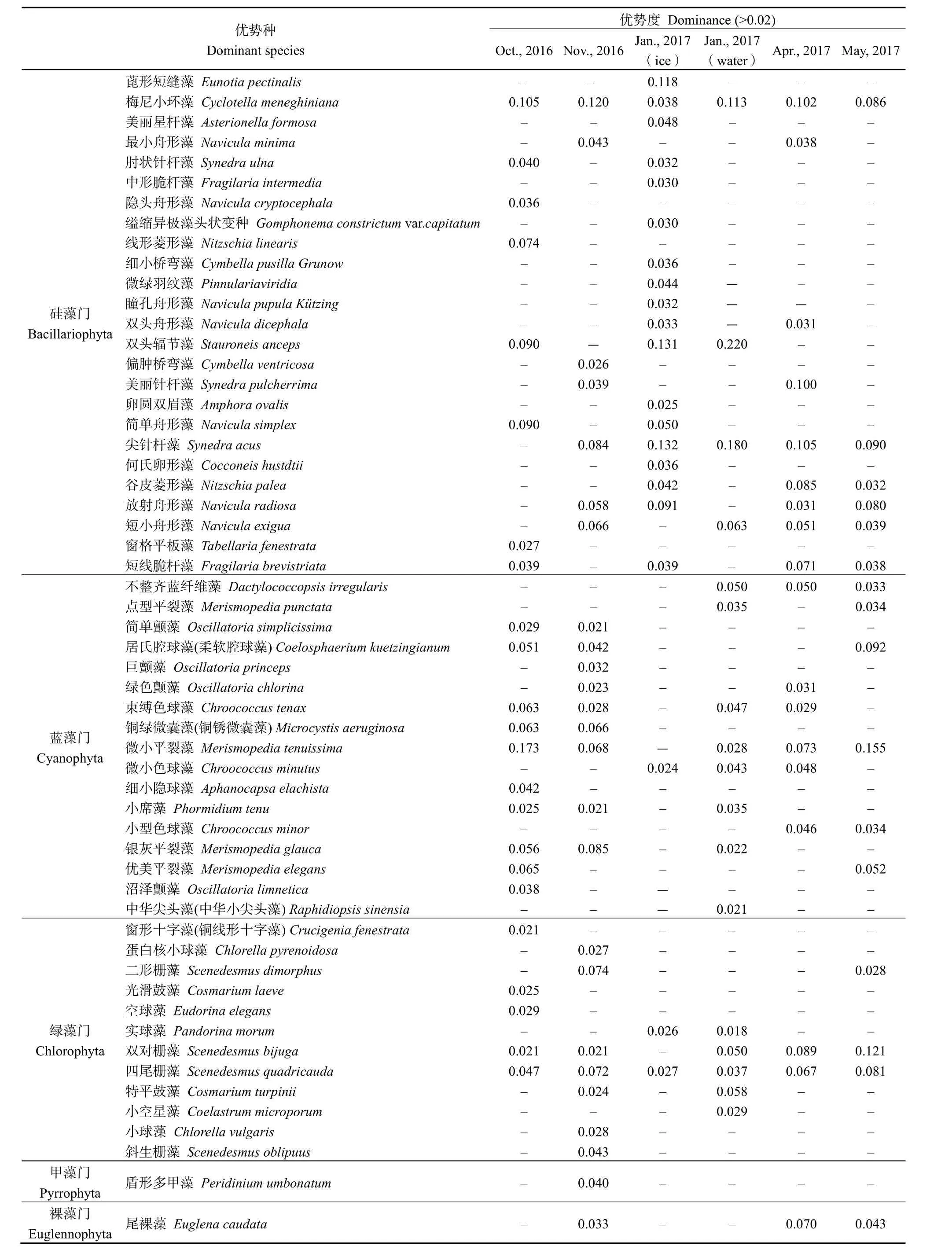
表1 乌梁素海浮游植物优势种名录Table 1 List of dominant species of phytoplankton in Wuliangsuhai
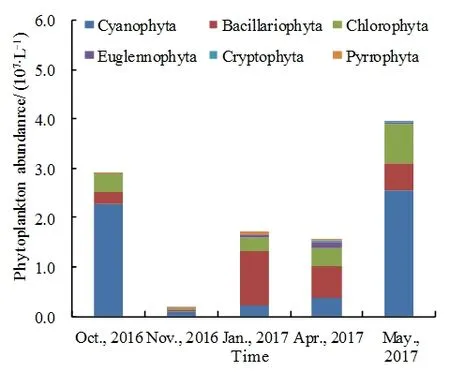
图4 乌梁素海浮游植物各门丰度随时间变化Fig. 4 Abundance of phytoplankton in Wuliangsuhai over time
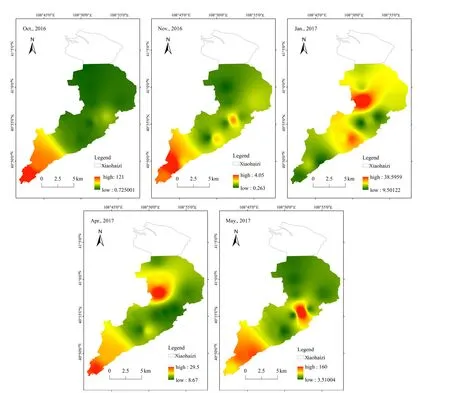
图5 乌梁素海浮游植物丰度空间变化Fig. 5 Spatial variation process of phytoplankton abundance in Wuliangsuhai
各月浮游植物丰度的空间差异较大(图5),10月表现为南部明显高于北部,南部明水区的采样点(二点、海壕、河口)形成密集区,其高值区主要分布蓝藻门的微小平裂藻(M. tenuissima)、优美平裂藻(M. elegans)以及细小隐球藻(A. elachista)等;11月也以南部明水区(海壕、河口)浮游植物丰度最高,其高值区主要分布有硅藻门的尖针杆藻(S. acus),蓝藻门的微小平裂藻(M. tenuissima)、银灰平裂藻(M. glauca)、铜绿微囊藻(M.aeruginosa)等;冰封期 1月浮游植物丰度以湖区北部较高,而硅藻基本遍布全湖,硅藻丰度占1月总体浮游植物丰度的63.02%,高值区主要分布有双头辐节藻(S.anceps)、尖针杆藻(S.acus)、蓖形短缝藻(E.factinalis)、放射舟形藻(N.radiosa)等;4月湖区北部以及南部浮游植物丰度有两个密集区,其高值区主要分布有蓝藻门的微小平裂藻(M.tenuissima),绿藻门的双对栅藻(S.bijuga)、四尾栅藻(S.quadricauda)等;5月湖区北部以及南部浮游植物丰度也有两个较为密集的区域,高值区主要分布有蓝藻门的居氏腔球藻(C.kuetzingianum)、微小平裂藻(M.tenuissima),绿藻门的双对栅藻(S.bijuga)、四尾栅藻(S.quadricauda),硅藻门的尖针杆藻(S.acus)、梅尼小环藻(C.meneghiniana)。
2.4 浮游植物多样性分析
多样性指数为浮游植物群落特征的重要参数,反映浮游植物群落结构的变化。从各月多样性指数均值来看(表 2),乌梁素海浮游植物 Shannon-Wiener多样性指数H以4月最高(3.59),1月最低(2.89);Margalef丰富度指数D以10月最高(4.66),1月最低(3.20);Pielou均匀度指数J在冰封期 1月较高(0.85),10月、5月较低(均为0.78)。
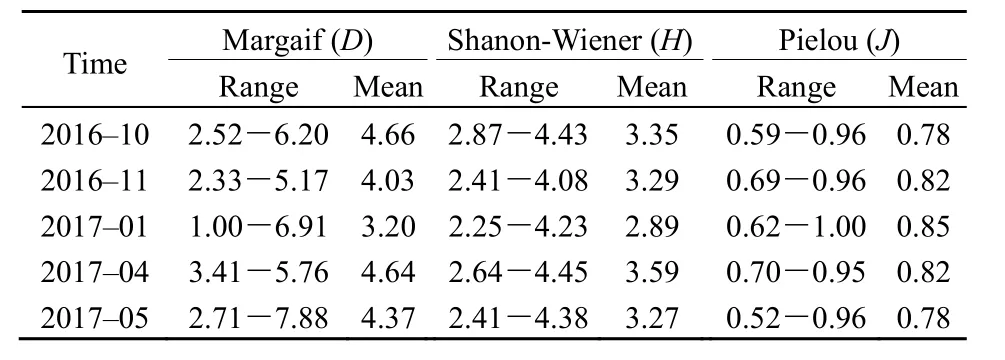
表2 乌梁素海浮游植物多样性指数Table 2 Phytoplankton diversity index of Wuliangsuhai
2.5 浮游植物与环境因子的RDA分析
为探明影响乌梁素海冻融前后浮游植物群落结构的环境因子,需先对物种数据进行非约束性分析——去趋势对应分析(DCA),若最大排序轴的长度大于 4,选择单峰模式模型;若小于 3,则选择线性模式;若介于 3-4之间,则单峰模型和线性模型均可(Temponeras et al.,2000)。通过对乌梁素海浮游植物细胞丰度与环境因子的多元关系进行去趋势对应分析,得到藻类细胞的最大梯度值为0.862<3,因此选择线性模型冗余分析(RDA)。在进行冗余分析(Borcard et al.,1992)时,选取环境指标水深(WD)、温度(t)、透明度(SD)、电导率(EC)、pH、溶解氧(DO)、氧化还原电位(ORP)、盐度(SAT)、总溶解性固体(TDS)、总氮(TN)、氨氮(NH4+-N)、硝态氮(NO3--N)、亚硝态氮(NO2--N)、总磷(TP)、化学需氧量(COD)、悬浮物(SS)与叶绿素a(Chl-a)与浮游植物月均丰度进行分析,每个环境因子的重要性和显著性采用Monte-Carlo假设进行检验(李红等,2014;Wang et al.,2014),以P<0.05筛选出对环境影响显著的因子(李艳利等,2015),分别为t(0.061)、SD(0.002)、EC(0.034)、pH(0.044)、DO(0.028)、TDS(0.026)、TN(0.044)、NH4+-N(0.001)、TP(0.001),然后对经过筛选的42种浮游植物(表3)及9个环境因子进行RDA分析。

表3 乌梁素海浮游植物种类代码Table 3 Species code of phytoplankton in Wuliangsuhai
RDA分析结果表明(表4),所选的9个环境因子共解释66.4%的物种变化,第1和第2排序轴分别贡献了39.6%和26.7%。在浮游植物物种与环境因子的排序图中(图6),箭头连线的长度表示环境因子与浮游植物群落分布相关程度的大小,连线越长,相关性越大,连线越短,相关性越小(张金屯,2011)。图中箭头长度从大到小依次为NH4+-N、TP、T、TDS、DO和SD,也相应代表了对浮游植物群落影响的重要程度。环境因子与前两个排序轴的相关系数见表 5,与轴 1呈显著正相关的有NH4+-N和pH,呈显著负相关的为 TP,NH4+-N、TP和pH箭头明显长于其他环境因子,主导着轴1方向物种的变化;与轴2呈显著正相关的有DO和SD,呈显著负相关的有t和TDS,其中SD和DO箭头明显长于其他环境因子,主导着轴2方向物种的变化。
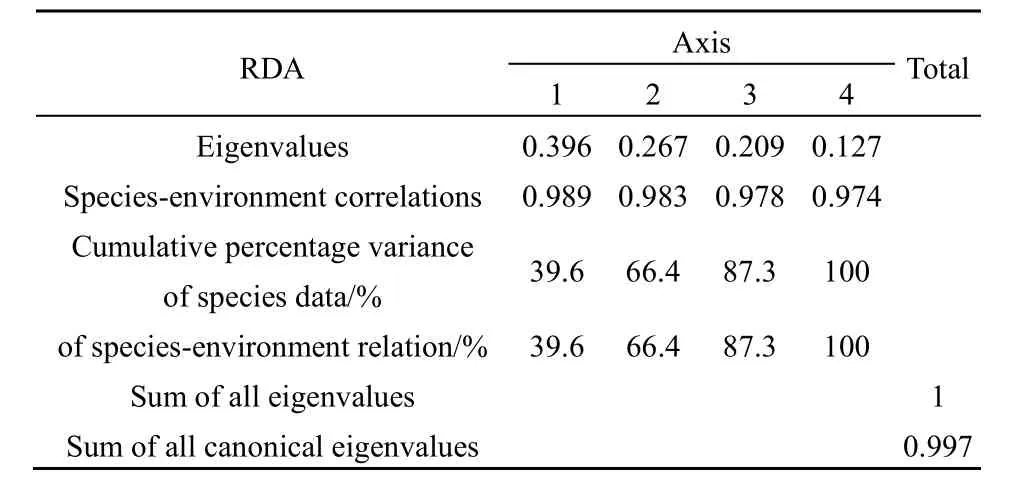
表4 浮游植物与环境因子的RDA分析结果Table 4 RDA analysis results of phytoplankton and environmental factors
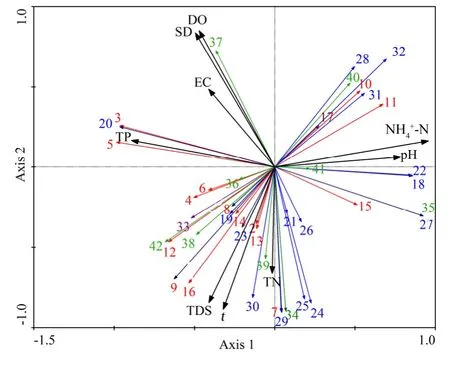
图6 浮游植物物种与环境因子的RDA分析Fig. 6 RDA analysis of phytoplankton species and environmental factors
3 讨论
3.1 乌梁素海冻融前后浮游植物群落结构特征
浮游植物种群结构结冰前蓝藻占优势,随着冰封期的到来,蓝藻占比渐少,硅藻占比渐多,融冰后蓝藻占比渐多,硅藻占比渐少。乌梁素海冰封期1月水体和冰体内浮游植物种类组成以硅藻门占绝大多数,而非冰封期浮游植种类组成多为蓝藻门、绿藻门占优势,表明非冰封期和冰封期水体浮游植物群落结构不同。硅藻生态位较宽,硅藻能够在低温条件下通过减少细胞中水分和增加细胞中糖类、脂肪等物质增强抵抗力,使其在低温条件下相比其他门类浮游植物更具有竞争力(李兴等,2017)。乌梁素海结冰前、融冰后南部明水区浮游植物大量聚集,因为这片区域的水域广阔、流速和缓、水动力不足以及营养盐浓度较高,这些条件均促使浮游植物大量聚集繁衍;而冰封期1月浮游植物丰度在湖区北部入湖口附近较高,由于冰封期1月入湖水量很少,没有农田退水的补给且水体结冰,且因冰盖的阻隔作用,水体流动性较小,生活污水与工业废水不易在全湖扩散。
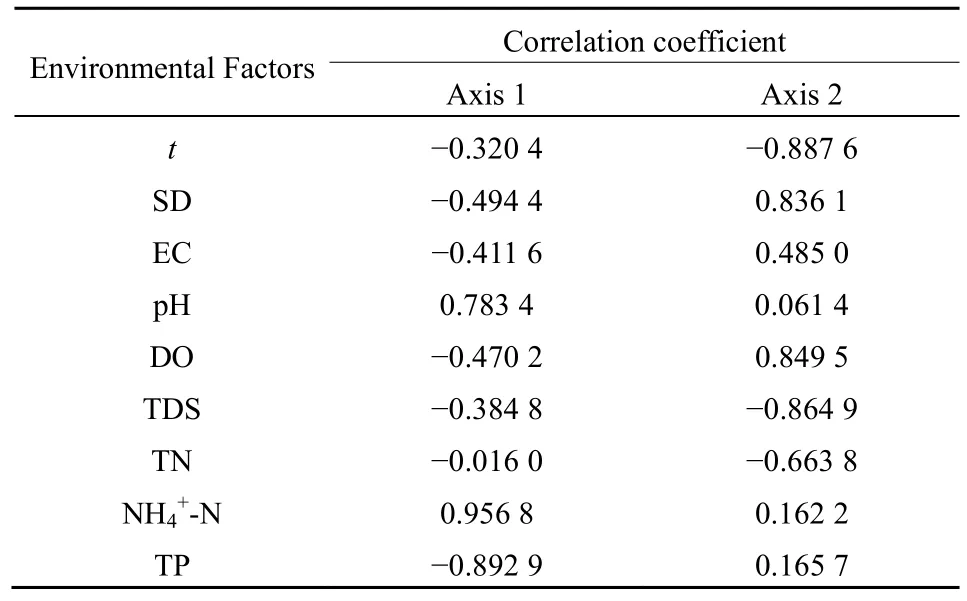
表5 环境因子与RDA主排序轴的相关系数Table 5 Correlation coefficients between environmental factors and RDA main sorting axes
3.2 乌梁素海冻融前后水质评价
利用指示种评价水质情况可知,优势种中优势度较大的微小平裂藻(M.tenuissima)适合于富营养型(α-ms)型水体,而梅尼小环藻(C.meneghiniana)以及尖针杆藻(S.acus)适合于富营养型(β-α-ms)型水体(沈韫芬等,1995),从这些优势种指示情况可知乌梁素海冻融前后月份水体处于富营养化水平。根据朱旭军等(2013)研究,Margalef丰富度指数D>5,水质清洁;D>4,寡污型;D>3,β中污型;D<3,α中污-重污型。乌梁素海冻融前后不同样点的Margalef 丰富度指数表明,除冰封期1月处于β中污型外(D>3),结冰前、融冰后各月皆指示为寡污型(D>4),丰富度指数表明乌梁素海冻融前后月份水体污染状况总体较轻。以Pielou均匀度指数J>0.3为标准(郭沛涌等,1997),则乌梁素海冻融前后浮游植物分布整体较均匀稳定。浮游植物多样性反映了群落稳定性,优势种种类和数量与多样性变化密切相关,若浮游植物群落优势种较多且各优势种优势度不高,那么浮游植物种群多样性就高,则这个浮游植物群落较复杂、稳定(况琪军等,2005)。本研究中,结冰前10月、11月与融冰后4月、5月优势种种类很多,但无优势性特别突出的种类,因此当季的Shannon-Wiener多样性指数H值、Margalef丰富度指数D值以及Pielou均匀度指数J值都较高。而冰封期1月虽各门优势种最多,但冬季1月温度比较低,喜温热环境的蓝藻和绿藻种类较少,硅藻门类优势种优势度显著,故 1月份的 Shannon-Wiener多样性指数 H值、Margalef丰富度指数D值均值变小。由上述分析可知,乌梁素海冻融前后月份水体污染状况较轻,浮游植物群落结构总体较均匀稳定,但冰封期1月水体浮游植物硅藻门占比很大,群落结构相对简单。
3.3 环境因子对乌梁素海冻融前后浮游植物群落结构的影响
浮游植物群落格局受多种环境因子的影响,如温度、营养盐、盐度、水动力条件以及浮游动物摄食等。通常地理位置不同,水体的生物以及非生物过程差别较大,从而导致浮游植物群落结构因子不同。侯伟等(2011)研究表明,苍村和赤石迳水库浮游植物群落主要受水深和透明度影响;沈会涛等(2008)研究得出影响白洋淀浮游植物群落的主要因素为pH值和总磷;李德亮等(2012)研究得出水深、水温、透明度、总磷、氧化还原电位以及电导率为影响大通湖浮游植物群落格局的主要因子。根据 RDA分析结果,影响乌梁素海冻融前后浮游植物群落结构变化的主要因子为:NH4+-N、TP、t、TDS、DO和SD。氮、磷是浮游植物生长繁殖的重要营养盐,也是常见的限制性营养元素,藻类优先同化吸收利用还原态氨(铵态氮),这与RDA分析结果一致。Parinet et al.(2004)认为,当N/P值大于7.2时,磷为限制性因素,本研究中,乌梁素海冻融前后月份 TN/TP 分别为 59.9、26.9、20.5、42.3、15.8,表明磷为乌梁素海该段时间内藻类增长的限制性因素。水温为影响浮游植物群落分布的关键因子(熊莲等,2016),硅、绿、蓝藻多聚集于第三象限,与水温呈正相关,浮游植物的生长需要在一定光照和温度下进行,不同种类浮游植物适宜生长的温度不同,RDA分析中,硅藻门在4个象限均有分布,说明其对乌梁素海冻融前后环境条件有较好适应性。透明度会直接影响浮游植物和其他水生生物的生存,总溶解性固体(TDS)减小,透明度增加,浮游植物丰度和生物量逐渐减小。水体中浮游植物与DO浓度密切相关,浮游植物进行光合作用释放氧气,使水域溶解氧浓度升高,浮游植物死亡降解也需要消耗溶解氧。
4 结论
(1)调查期间,在乌梁素海观察到的浮游植物共计6门69属120种,种类组成以硅藻为主;浮游植物种群结构结冰前以蓝藻占优势;随着冰封期的到来,蓝藻占比渐少,硅藻占比渐多;融冰后蓝藻占比渐多,硅藻占比渐少。乌梁素海冰封期前后月份南部明水区浮游植物聚集,冰封期1月浮游植物丰度在湖区北部入湖口附近点位较高。
(2)根据优势种指示情况可知,乌梁素海冻融前后水体处于富营养化状态;丰富度指数反映水体总体污染状况较轻,均匀度指数反映浮游植物分布整体较均匀;冰封期1月水体浮游植物硅藻门占比很大,群落结构相对简单。
(3)RDA 排序揭示氨氮(NH4+-N)、总磷(TP)、温度(t)、总溶解性固体(TDS)、溶解氧(DO)和透明度(SD)为影响乌梁素海水体冻融前后浮游植物物种分布格局的主要环境因子。
