不同放牧强度下荒漠草原土壤氮矿化季节性动态研究
2019-05-31单玉梅温超常虹张璞进晔薷罕木兰王常慧黄建辉白永飞孙海莲陈海军
单玉梅 ,温超 ,常虹 ,张璞进 ,晔薷罕 ,木兰 ,王常慧,黄建辉,白永飞,孙海莲 ,陈海军
1. 内蒙古自治区农牧业科学院,内蒙古 呼和浩特 010031;2. 中国科学院内蒙古草业研究中心,内蒙古 呼和浩特 010031;3. 中国科学院植物研究所/植被与环境变化国家重点实验室,北京,100093;4. 内蒙古自治区生物技术研究院,内蒙古 呼和浩特 010010
荒漠草原是草原向荒漠过渡的主要植被类型,也是内蒙古草原的重要组成部分,是中国北部边疆的生态安全屏障。作为草原畜牧业主要生产资料地,荒漠草原处于极特殊的生态地域,生态系统稳定性差,植被种类比较贫乏,以强旱生的小半灌木或矮禾草为主,草层低矮、群落结构单一(马治华等,2007)。放牧是天然草地最主要的利用方式之一,也是一个典型的人为干扰因素(陈利顶等,2000)。因此,荒漠草原生态学过程容易受到自然因素和人为因素的双重干扰(李德新,1990)。放牧家畜主要通过采食、践踏和排泄粪便3种主要形式影响草地营养物质的循环,使草地土壤养分发生变化,土壤养分之间也是相互作用,相互制约(An et al.,2015;Risch et al.,2015;Jiang et al.,2016;Wei et al.,2017;Wang et al.,2016)。
氮素不仅是大气圈中含量最丰富的元素,也是各种植物生长和发育所需的大量营养元素之一,氮素的有效性是草地生产力的重要影响因子之一,氮矿化速率的高低反映了土壤的供氮能力,其影响因素可归结为生物因子(土壤动物、土壤微生物和植物种类等)、非生物因子(土壤温度,土壤含水量,土壤理化性状等)和人类活动(主要包括放牧、割草、施肥和火烧等)(王常慧等,2004;朱志成等,2017;杨浩等,2017;罗亲普等,2016;Shan et al.,2011;Wang et al.,2006;2011;Petrie et al.,2016)。
内蒙古荒漠草原的气候属于水热同期变化,土壤温度和含水量存在季节变化,是影响土壤氮矿化的重要环境因子,因此,研究放牧强度对土壤氮矿化速率影响的季节性变化,对于了解放牧强度对氮素可利用性的影响具有重要意义。本研究依托长期放牧控制试验平台,研究荒漠草原土壤净氮矿化速率的季节性变化对放牧强度的响应,结合植物群落、气候因子和土壤理化性质的测定,综合分析土壤无机氮库、净氮矿化速率和生态系统功能之间的内在联系,解释放牧强度对土壤氮矿化过程的影响机理,为合理利用草地资源和荒漠草地生态系统的恢复提供理论依据。
1 试验区概况
试验地位于内蒙古乌兰察布市四子王旗王府一队,地理位置为 41°47′17″N 和 111°53′46″E,平均海拔在1450 m左右,气候属于典型的大陆性季风气候,春季干旱,大风频繁,夏季炎热,降雨量集中期为 5-8月,多年平均降雨量为 280 mm,≥10 ℃积温为1900-2500 ℃,平均无霜期为120 d,平均温度最高的月份是 7月。土壤类型为淡栗钙土,距地表50-60 cm处多出现白色钙积层。草地类型为短花针茅+冷蒿+无芒隐子草的荒漠草原,植被草层比较低矮,一般在8 cm左右,植被较稀疏,盖度在15%-20%之间,群落主要由20多种植物组成,建群种为短花针茅(Stipa breviflora),优势种为冷蒿(Artemisia frigid)和无芒隐子草(Cleistogenes songorica)。
2 材料与方法
2.1 试验设计
实验区设在内蒙古农牧业科学院综合试验示范中心的长期放牧控制试验平台。该放牧试验平台已有连续 10年的放牧历史。采用随机区组设计,将试验区分成3个区组,为3个重复。每个区组内设有1个无牧区和3个放牧区,其中无牧区作为对照(CK)、,放牧区分别设3个放牧强度,分为轻度放牧(LG)、中度放牧(MG)和重度放牧(HG),4个牧区载畜率放牧强度分别为0、0.93、1.82、2.71只羊·hm-2。放牧时间为6月初到11月底,共计6个月,白天将供试羊只赶入相应试验围栏内让其自由采食,晚上相同时间归圈休息。试验期间保证羊只充足的饮水量,定期补盐,且其他放牧管理方式均一致。冬季来临,供试羊只则实行圈养的管理方式,以秸秆、青干草和玉米作为主要补饲草料。
2.2 测定方法
2.2.1 土壤无机氮含量和净氮矿化速率的测定
采用野外原位顶盖 PVC管法测定土壤净氮矿化速率。具体方法为:首先去除地表的植物和凋落物,将一对PVC管(内径5 cm×长12 cm)插入地下10 cm,其中一个PVC管顶端用透气不透水的无菌培养封口膜封口,野外培养 30 d。将另一 PVC管中的土壤装入封口袋中,带回实验室。将取回的土样过2 mm筛,充分混匀,分装成2份,一份保持为鲜土,用于测定初始无机氮含量;另一份进行风干处理,用于测定土壤的pH值、有机质、全氮及全磷等含量。
试验从2013年5月开始,2014年4月结束,期间共取样8次,时间间隔根据季节性的气候因子变化划分为3个取样阶段:(1)生长季初期,即2013年5-6月;(2)生长季盛期,即2013年7-9月;(3)非生长季,即2013年10月-2014年4月。生长季初期的取样时间为2013年5月初和6月初,生长季盛期是每月月初取样1次,野外培养时间约为30 d,非生长季的取样时间为2013年10月、11月和2014年4月,野外培养时间为30-60 d。
2.2.2 数据处理
净氮矿化速率是单位时间内土壤无机氮(NH4+-N+NO3--N)的变化量,即培养后土壤无机氮含量与培养前土壤无机氮含量的差值。累积氮矿化量等于整个取样期间每个培养时期净氮矿化量的加和。净氮矿化速率和硝化速率的计算公式如下:

式中,Δt表示培养时间;ti表示培养前的时间;ti+1表示培养后的时间;c[NH4+-N]i和 c[NH4+-N]i+1分别表示培养前后铵态氮的浓度;c[NO3--N]i和c[NO3--N]i+1分别表示培养前后硝态氮的浓度;Aamm、Anit和 Amin分别表示培养期间铵态氮、硝态氮和总无机氮(铵态氮和硝态氮)的累积量;Rmin和Rnit分别表示净氮矿化和净硝化速率。
2.2.3 数据分析
数据分析运用SAS 9.0软件,(1)采用Repeated Measures ANOVA方法检验不同季节(非生长季、生长季初期和生长季盛期)放牧强度对土壤温度、含水量、无机氮含量、净氮矿化和净硝化速率的影响。(2)采用One-way ANOVA方法,分别比较每一季节不同放牧强度对土壤净氮矿化速率和累积氮矿化的差异,并采用 Duncan′s方法进行多重比较分析。(3)土壤无机氮含量、净氮矿化速率与土壤温度、土壤含水量之间的关系采用Pearson's相关分析检验。
3 结果分析
3.1 不同放牧强度下土壤无机氮含量的季节动态
方差分析结果表明,整个年度放牧强度对土壤无机氮含量无显著影响(P>0.05),但取样时间对土壤无机氮含量有显著影响(P<0.05),放牧强度和取样时间的交互作用仅对生长季初期的铵态氮含量有显著影响(P<0.05)(表1)。
整体而言,土壤铵态氮含量在生长季初期(2013年5-6月)逐渐增加,生长季盛期先降低后升高,9月在对照区达到峰值(6.109 g·kg-1),在非生长季(2013年10月-2014年4月)呈逐渐减少趋势(图1)。

图1 不同放牧强度下土壤无机氮含量的季节动态变化Fig. 1 Patterns of soil NH4+-N, NO3--N, and total inorganic N concentrations across seasons under different grazing intensities
土壤硝态氮含量在生长季初期和非生长季呈现出与铵态氮含量相反的季节变化趋势,生长季盛期呈现出与铵态氮含量变化相似的模式,其峰值(11.299 g·kg-1)出现在9月的LG区(图1)。土壤总无机氮含量变化与硝态氮含量变化一致的趋势,峰值(17.318 g·kg-1)也出现在9月的LG区。生长季初期土壤总无机氮含量高于生长季盛期和非生长季(图1)。
3.2 土壤净氮矿化作用与净硝化作用
整个年度放牧强度对土壤净氮矿化速率和净硝化速率均无显著影响(P>0.05),取样时间显著影响土壤净氮矿化和净硝化速率(P<0.05),其他季节均无显著影响(P>0.05)(表1)。
在生长季初期,随放牧强度增加土壤净氮矿化速率逐渐降低,并且在MG和HG出现了氮固持现象,HG固持量最大;对照区和LG区的土壤净氮矿化速率分别为 0.010 g·kg-1·d-1和 0.003 g·kg-1·d-1,MG 和 HG 区的分别为-0.003 g·kg-1·d-1和-0.010 g·kg-1·d-1(图 2A)。土壤净硝化速率也随放牧强度增加而逐渐降低,对照区的净硝化速率为 0.033 g·kg-1·d-1,放牧区该指标的变幅为0.002-0.043 g·kg-1·d-1,HG 区显著最低(P<0.05)(图2B)。
在生长季盛期,随放牧强度增加土壤净氮矿化速率和净硝化速率有增加趋势,对照区土壤净氮矿化速率为 0.060 g·kg-1·d-1,LG 区的净氮矿化速率为0.146 g·kg-1·d-1,比对照区高 145%;土壤净硝化速率的峰值出现在 LG 区(0.213 g·kg-1·d-1),比 CK区(0.166 g·kg-1·d-1)高 30%(图 2A、B)。
在非生长季,随放牧强度增加土壤净氮矿化速率和净硝化速率有降低的趋势,土壤净氮矿化速率变幅为 0.075(MG)-0.178 g·kg-1·d-1(CK),土壤净硝化速率的均值变幅为 0.070(MG)-0.162 g·kg-1·d-1(CK),峰值出现在 CK 区,净硝化速率显著高于MG区(P<0.05)(图2A、B)。

表1 放牧强度对土壤无机氮含量、净氮矿化和硝化速率季节变化影响的方差分析结果Table 1 F values of repeated measures analysis of variance for soil inorganic N concentrations (NH4+-N, NO3--N, and NH4+-N+NO3--N) at the start of each incubation period and daily net N mineralization (Rmin) and nitrification (Rnit) rates over different seasons using grazing intensity (GI), sampling time (Month),and GI×Month as fixed-effects

图2 不同季节土壤氮矿化速率和硝化速率对放牧强度的响应模式Fig. 2 Effects of grazing on soil net N mineralization and nitrification rates over the non-growing season, early growing season and peak growing season
3.3 不同季节的累积氮矿化量与硝化量
生长季初期,随放牧强度增加土壤累积氮矿化量和硝化量逐渐减少,峰值均出现在 LG,显著高于MG和HG(P<0.05),并且MG和HG均出现氮固持现象,最大的累积氮固持出现在HG区(-0.815 g·kg-1)(图 3A、B)。
生长旺季,随放牧强度的增加土壤累积氮矿化量和硝化量逐渐增加,HG的累积氮矿化量最高,为1.583 g·kg-1,显著高于 LG和MG区(P<0.05),并且LG和MG区均出现氮固持现象;对照区累积硝化量显著高于LG和MG处理区(P<0.05),分别高出3倍和2倍(图3C、D)。
非生长季,随放牧强度增加土壤累积氮矿化量和硝化量呈现出先降低后升高的趋势。与对照相比,放牧显著降低了累积氮矿化量,特别是在MG区,降幅达142%(P<0.05)(图3E、F)。
从全年来看,放牧降低了土壤累积氮矿化量和硝化量,尤其是累积硝化量显著低于对照区(P<0.05)(图 4)。
3.4 土壤无机氮和净氮矿化对环境因子和土壤性状的季节响应
放牧强度、取样时间及其交互作用显著影响土壤温度和含水量(P<0.0001)。在生长季初期,土壤温度与土壤含水量具有相似的变化趋势,但在生长季盛期和非生长季,它们的变化模式呈相反的趋势。相关分析结果也表明,土壤温度与土壤含水量之间呈显著的负相关关系(r=-0.374,P<0.0001),土壤含水量的峰值出现在非生长季的冻融期(3月)。与MG和HG区相比,CK和LG区具有较低的土壤温度和较高的土壤含水量(P<0.05)(图5)。

图3 不同季节土壤累积氮矿化量和硝化量对放牧强度的响应模式(生长季初期A和B,生长季盛期C和D,非生长季E和F)Fig. 3 Effects of grazing on cumulative net N mineralization (A, C, and E) and nitrification (B, D, and F) over the non-growing season, early growing season and peak growing season

图4 整个季节土壤累积氮矿化量和硝化量对放牧强度的响应模式Fig. 4 Effects of grazing intensity on cumulative net N mineralization and nitrification over the whole season

图5 不同放牧强度下土壤温度和土壤含水量的季节变化Fig. 5 Seasonal changes of soil temperature and soil moisture content at the start of in situ N mineralization assays under different grazing intensities
Pearson's相关分析表明,土壤温度和土壤含水量是调控土壤无机氮和净氮矿化季节动态的两个重要因素,但二者对无机氮库、净氮矿化和净硝化速率的影响随季节变化发生改变。在生长季初期,土壤温度与铵态氮含量呈显著正相关,与硝态氮和总无机氮含量呈显著负相关(P<0.05)。在生长季盛期,土壤温度与无机氮含量、净氮矿化速率和净硝化速率均呈显著正相关(P<0.05);与净氮矿化速率和净硝化速率均呈显著正相关关系(P<0.05)。在非生长季,土壤温度与铵态氮含量之间呈显著正相关关系,与硝态氮和总无机氮含量之间呈显著负相关关系(P<0.05);土壤含水量与铵态氮含量和矿化速率和硝化速率之间均呈显著负相关关系,与硝态氮和总无机氮含量之间呈现显著正相关关系(P<0.05)(表2)。
单因素方差分析结果表明,随放牧强度增加,土壤容重和硬度显著提高,地上净初级生产力逐渐降低,与CK区相比,LG、MG和HG分别减少了46.2%、64.4%和 76.6%,且各处理间差异显著(P<0.05)(图 6)。
4 讨论
4.1 放牧对土壤净氮矿化的影响
本研究发现随着气候因子(温度和降水)的季节性变化以及草原植物生长的节律变化,放牧对土壤净氮矿化作用的影响存在着显著的季节性差异。在生长季初期放牧抑制了土壤净氮矿化作用,主要归因于长期的干旱抑制了植物生长和土壤微生物的活性,进一步抑制了土壤净氮矿化。本研究结果表明,表层土壤温度随着放牧强度的增加而上升,土壤含水量随着放牧强度的增加而显著降低,且相关分析结果也发现土壤无机氮含量与土壤温度呈显著负相关、与土壤含水量呈显著正相关。Shan et al.(2011)在典型草原发现,随放牧强度的增加,土壤温度和土壤含水量具有相反的变化趋势,与本研究结果一致。
在生长季盛期,放牧促进了土壤净氮矿化作用,其中LG区的净氮矿化速率和净硝化速率均较高,说明适度放牧有利于土壤净氮矿化的发生,这与Shan et al.(2011)、Austrheim et al.(2014)、Wang et al.(2016)、Yan et al.(2016)和Zhou et al.(2017)的研究结果一致。放牧,尤其是轻度放牧,对土壤净氮矿化的促进作用机制是放牧改变了植物和土壤微生物群落结构(Risch et al.,2015;Lima et al.,2016),通过刺激植物根际微生物活动(Devi et al.,2014)和家畜粪尿的分解(An et al.,2015;Wang et al.,2016)加强净氮矿化作用,家畜活动促使碳分泌物增多,使土壤矿化能力增强,有利于轻度放牧区氮素的转化(Zhang et al.,2008;Bardgett et al.,2003)。另外,本研究发现,放牧区的植被返青时间比对照区提前 2-3周,植物生理活动增加了根际碳分泌物排放,从而促进土壤净氮矿化。9月份铵态氮和硝态氮含量均较高,也可能是加剧生长季盛期土壤净氮矿化作用的原因,因为有研究表明,草地土壤净氮矿化速率受硝化作用的影响较大,硝化速率与净氮矿化速率呈显著正相关(韩梦琪等,2017)。

表2 土壤无机氮含量、净氮矿化和净硝化速率、净氮矿化累积量和净硝化累积量与土壤温度、土壤含水量之间的相关关系Table 2 Pearson's correlation coefficients (r) and P values of soil NH4+-N, NO3--N and total inorganic N contents, net N mineralization (Rmin) and nitrification(Rnit) rates, cumulative net N mineralization (Cmin) and nitrification (Cnit) with soil temperature and moisture for each season across all grazing intensities
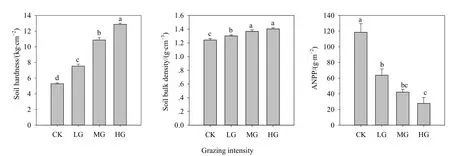
图6 不同放牧强度下土壤理化性质和地上净初级生产力的变化Fig. 6 Variations of soil hardness, soil bulk density and aboveground net primary production (ANPP) under different grazing intensities
非生长季放牧抑制了土壤净氮矿化作用,对照区土壤净氮矿化速率显著高于放牧小区。Shan et al.(2011)发现随放牧强度增加,地上净初级生产力和植被覆盖度降低,放牧区的积雪厚度降低,而对照区地上立枯较多,有较高的积雪厚度,有利于土壤净氮矿化的发生(Schimel et al.,2004;刘琳等,2011),并且改变植物可利用氮的数量和有效性(Wanet al.,2011)。另外,对照区积雪厚度较高,导致土壤温度较高,相反,放牧区积雪厚度较低导致土壤温度较低(Shan et al.,2011),抑制了土壤净氮矿化作用(Schimel et al.,2004;刘琳等,2011);而非生长季的冻融现象对于土壤氮转化过程也有很重要的影响(Schimel et al.,2004;Shan et al.,2011;赵媛等,2017;Zhao et al.,2010;Kielland et al.,2006)。
4.2 净氮矿化作用的季节模式对土壤温度和含水量的响应
随着全年土壤温度和土壤含水量的季节性变化,土壤无机氮库、净氮矿化速率也发生相应的变化。研究结果显示,取样时间显著影响着土壤铵态氮、硝态氮和总无机氮含量以及净氮矿化和净硝化速率,Pearson's相关分析也揭示了影响土壤无机氮和净氮矿化季节动态模式的两个主要环境因素是土壤温度和含水量。荒漠草原生态系统的这一结果与前人在典型草原(Shan et al.,2011;朱志成等,2017;Wang et al.,2006)、高寒草甸草原(武丹丹等,2016)和森林生态系统(Kielland et al.,2006;李志杰等,2017)的研究结果一致。
在生长季初期,土壤无机氮含量与土壤温度呈显著负相关,而与土壤含水量无显著相关关系,土壤净氮矿化速率与土壤温度和含水量均无显著相关关系,与Wang et al.(2006)在典型草原生态系统的研究结果(土壤净氮矿化速率与温度和水分均正相关)不同。原因可能是荒漠草原的土壤养分条件相对于典型草原生态系统较差(朱剑兴等,2013),土壤温度随季节的深入不断升高,但该期间降水十分稀少,呈现土壤水分与温度变化的不一致性,导致土壤严重干旱,土壤含水量的作用不明显,微生物作用也显著降低,而土壤水分是影响微生物活性、加快土壤有机氮分解的重要因素之一(Cregger et al.,2014;Delgado-Baquerizo et al.,2014)。在生长季盛期,土壤净氮矿化速率和硝化速率与土壤温度和含水量均呈现极显著的正相关关系,与生长季初期试验结果不同,原因如下:7-9月是降雨的高峰期,也是荒漠草原植被和土壤状况最佳的时期,因此土壤净氮矿化速率受温度和水分的影响规律和典型草原一致;其次土壤氮矿化具有相对较强的季节性,季节的更替直接影响着土壤含水量和土壤温度,土壤微生物活动受土壤温度和土壤水分含量的影响,进一步影响土壤净氮矿化速率的变化(Shan et al.,2011;朱志成等,2017;Wang et al.,2006,2011;Devi et al.,2014;Cregger et al.,2014;Delgad-Baquerizo et al.,2014)。土壤含水量与土壤无机氮含量呈显著正相关,与净氮矿化速率和硝化速率均呈显著负相关,与韩梦琪等(2017)在荒漠草原生态系统的研究得出的结果一致。土壤无机氮的重要来源是土壤可溶性有机氮,而冻融可以促进土壤可溶性有机氮的释放(陈哲等,2016),但是,冻融也会导致土壤微生物活性降低甚至死亡,将直接影响土壤净氮矿化速率(王连峰等,2007)。
在干旱半干旱草原,水和氮的耦合不仅对诸如土壤氮素转换的养分循环过程有显著的影响,同时对其他生态系统功能起着重要的作用(Bai et al.,2004,2010)。本研究发现,放牧对土壤无机氮含量和净氮矿化作用的影响与地上净初级生产力的变化密切相关,高强度放牧抑制了土壤氮矿化过程,这表明氮素的分配模式也受放牧强度的制约(An et al.,2015;Jiang et al.,2016;Shan et al.,2011)。在较高的放牧强度下,植物需要分配较多的氮素给地上组织,以弥补家畜造成的地上部分的氮损失,重新构建光合组织,而地上组织含氮量增加即可形成较多的凋落物,进而促进凋落物分解(Semmartin et al.,2008),加快氮素循环。但是,根部氮素的减少会阻碍微生物活动产生负反馈,抑制土壤净氮矿化。本结果进一步说明了放牧对土壤无机氮库与净氮矿化作用的影响与生态系统功能变化之间存在着密切关系,需要继续进行深入研究。
5 结论
从全年来看,放牧强度对土壤无机氮含量、土壤净氮矿化速率和净硝化速率均无显著影响(P>0.05),但取样时间对土壤无机氮含量、土壤净氮矿化和净硝化速率有显著影响(P<0.05),放牧强度和取样时间的交互作用仅对生长季初期的铵态氮含量有显著影响(P<0.05),其他季节均无显著影响(P>0.05);放牧降低了土壤累积氮矿化量和硝化量,尤其是累积硝化量显著低于对照区(P<0.05)。随放牧强度增加,土壤容重和硬度显著提高,地上净初级生产力逐渐降低,且各处理间差异显著(P<0.05)。放牧强度、取样时间及其交互作用显著影响土壤温度和含水量(P<0.0001)。在生长季初期,土壤温度与土壤含水量具有相似的变化趋势,但在生长季盛期和非生长季,它们的变化模式呈相反的趋势。土壤温度与土壤含水量之间呈显著的负相关关系,土壤温度和土壤含水量是调控土壤无机氮和净氮矿化季节动态的两个重要因素,但二者对无机氮库、净氮矿化和净硝化速率的影响随季节变化而变化。
