喀斯特山地草地群落多样性海拔特征及土壤理化性质特征
2019-05-31金章利刘高鹏周明涛许文年
金章利,刘高鹏,周明涛,许文年
1. 湖北三峡职业技术学院,湖北 宜昌 443000;2. 湖北金源绿通生态环境工程有限公司,湖北 宜昌 443000;3. 三峡大学土木与建筑学院,湖北 宜昌 443002
土壤是草地生态系统的重要组成部分,不仅为动物及微生物提供了赖以生存的栖息场所,也为植物提供必需的营养和水分,是各种物质能量转化的场所(权国玲等,2015;孙元丰等,2018)。其养分含量的高低不仅影响植物个体发育,更进一步决定着植物群落的类型、分布和动态(饶胜等,2015)。植被对土壤养分的效应是植物的吸收与固定、群落生物量的积累与分解等,使得土壤养分在时间和空间尺度上出现了各种动态变化过程(张雪峰等,2015),因此植被的土壤理化性质特征与植物群落的地上/地下生物量的大小、保存率和周转率等是分不开的(张继平等,2015)。土壤和植被具有互动效应,决定了土壤与植被总是处于不断的演化与发展之中(张继平等,2015)。大量研究结果表明,草地类型的变化可以引起许多自然因素和生态过程的变化,草地类型不同会影响土壤机械组成、土壤养分、植被特征等(李学斌等,2014)。
中国幅员辽阔,具有较为复杂的气候特点和地形地貌。就喀斯特地貌而言,中国分布面积最广,且主要分布于西南地区(李胜平等,2016)。该地区不仅地形复杂,且生态环境脆弱,仅略强于寒漠地区生态(武德传等,2016)。喀斯特山地景观存在巨大差异,具有独特的二元地貌特点,地上及地下相互联系、关系紧密,同时又相互制约,这也造就了典型差异的地上及地下生物群落分布(张红旗等,2017;许尔琪等,2016)。
中国喀斯特山地地形复杂、地势连绵,存在多样性明显的生物资源,同时体现出较大差异的生境高度,造就了植被分布的多样性,也形成了微生物分布的多样性(孙嘉曼等,2016)。贵州省睛隆县超过一半面积为喀斯特地形,该地区分布较多的是峡谷,崎岖的地表加之陡峭的山坡,在雨水及风力等作用下,地表侵蚀较为严重,土层较薄,蓄水能力较弱,土壤难以为植被生长提供充足养分,故其植被分布较稀疏,生长缓慢,植被生态受到很大的制约,整个局部生态缺乏稳定性和抗逆性,自我修复能力弱(尚二萍等,2017)。此外,近年来不合理的开发利用更加剧了该区域生态水平退化,受到该区域环境条件的影响,该区域不仅耕地少,同时形成了典型的石头多、土地少的特点,土壤侵蚀明显,石漠化现象较为明显,且短期内难以自我修复,整个生态环境较差(欧芷阳等,2015)。由于该地区脆弱的生态已经深深影响到当地的生活,越来越多的学者开展了相关的研究,不仅从土壤及植被的角度来探究该区域地貌特点,而且从耕种等方面开展相应研究(欧芷阳等,2015;尚二萍等,2017)。本研究在以往学者研究的基础上,采用经典统计分析与排序分析,连续3年研究了喀斯特山地草地群落多样性海拔特征及土壤理化性质特征,并探讨了群落多样性与土壤理化性质之间的关系,为喀斯特山地草地生态系统的深入研究和草地健康状况的客观评价提供依据。
1 材料与方法
1.1 研究区概况
研究区山高谷深坡陡,呈现典型的山峦起伏,并形成了较大差距的海拔,其最高、最低海拔相差近 1500 m。受所处位置的影响,该区域呈现明显的亚热带季风气候,雨热同季,具有明显的垂直分布性,山谷之处较为干热,而高原之处气温较低。该区域年降雨量1650 mm,年均气温15 ℃(图1),日照时间1500 h,无霜期320 d,降雨充沛且光热充足。然而,由于土壤土层较薄,土壤养分含量较低,持水性弱,难以充分满足植被生长,植被以山地草地为主。该区域自然灾害多发,岩溶等现象明显,喀斯特地貌特征显著(欧芷阳等,2015)。
1.2 试验设计
以贵州省睛隆县喀斯特山地草地(天然草地,未干扰)为研究对象(25°33′-26°11′N,105°01′-105°25′E,土地总面积 13.31×104hm2),分别沿海拔梯度布设样点,实验开始于2015年9月,分别在海拔 689 m(低海拔,优势种为金丝草Pogonatherum crinitum、矛叶荩草Arthraxon lanceolatus和画眉草Eragrostis pilosa等)、1021 m(中海拔,优势种为矛叶荩草Arthraxon lanceolatus、鼠尾粟Sporobolus fertilis、野青茅Deyeuxia arundinacea、紫荆泽兰Eupatorium adenophorum等)、1332 m(高海拔,优势种为矛叶荩草Arthraxonlanceolatus、扭黄茅Heteropogon contortus、三脉紫菀Aster ageratoides、香薷Elsholtzia ciliata)设置调查样点(表1),每个调查样点随机设置50个具有代表性的典型喀斯特山地草地植物群落特征的 100 cm×100 cm样方,以后连续3年重复采样,记录坡度、坡向、海拔和经纬度。
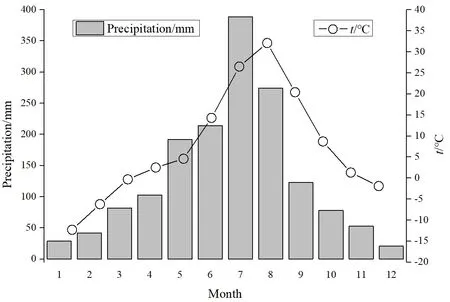
图1 研究区近3年平均气温和降水量月值变化Fig. 1 Changes of average temperature and precipitation in the study area in recent 3 years
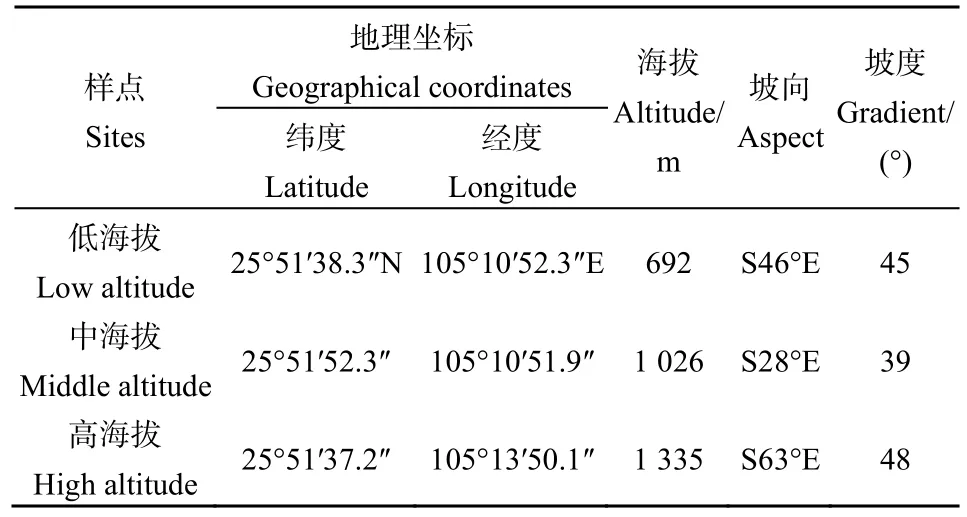
表1 长期定位样点描述Table 1 Description of long-term fixed sample point
1.3 生物量测定
在2015-2017年8月中旬,在上述每个样方进行植被调查,分别统计每个样方内的草地植物物种及每个种群的密度、盖度及高度,用于计算群落的物种多样性,具体如下:


式中,N为植物种种数。

式中,Pi为重要值;RD为相对密度;RS为相对优势度;RC为相对盖度。
植被调查完后,收获样方生物量,具体步骤如下:将100 cm×100 cm的样方分为4个25 cm×25 cm小样方,用剪刀齐地收割地上部,带回实验室杀青15 min后于65 ℃下烘至恒重,获得地上生物量(S);同时用铁锹挖取25 cm×25 cm小样方的所有植物根系(挖取深度为80 cm(,80 cm以下土层基本没有根系),去除死根(肉眼区分,黑色、空心、腐烂的根为死根),将挖取的根带回实验室,量取主根长,用清水冲洗干净后于65 ℃下烘至恒重,获得地下生物量(R);计算根冠比(R/S)。
1.4 土壤样品测定
采用四分法采集样方土壤,首先在各样方中采集1 kg土壤,取样深度最大为40 cm,将同一海拔土样混合均匀后带回实验室,剔除杂质后自然风干,待养分测定。使用元素分析仪测定全碳及氮含量,分别采用比色法、光度计法测定速效磷、钾含量(John et al.,2007)。
1.5 数据处理
运用Excel 2007和SPSS 18.0进行数据统计和方差检验,以平均值±标准误差表示(Mean±SE)。采用单因素方差分析(One-way ANOVA)检验不同海拔高度各指标的差异显著性;采用Pearson相关系数法检验各指标之间的相关性,所有原始数据进行对数转换;RDA经蒙特卡洛(Monte Carlo)检验(499次)后运用CANOCO 4.5分析土壤理化特征对植被多样性的响应。
2 结果与分析
2.1 不同海拔山地草地地上与地下生物量
由表2可知,低海拔草地根长为32.61-33.78 cm,中海拔草地为 38.96-39.77 cm,高海拔草地为 35.62-36.79 cm,大致表现为中海拔>高海拔>低海拔,不同海拔草地根长差异均显著(P<0.05);低海拔草地高度为56.98-58.12 cm,中海拔草地为61.45-65.99 cm,高海拔草地为59.32-63.07 cm,大致表现为中海拔>高海拔>低海拔,不同海拔草地高度差异均显著(P<0.05);低海拔草地地上生物量(AGB)为23.15-24.77 g·cm-2,中海拔草地为32.47-35.02 g·cm-2,高海拔草地为 28.77-30.14 g·cm-2,大致表现为中海拔>高海拔>低海拔,不同海拔草地 AGB差异均显著(P<0.05);低海拔草地地下生物量(BGB)为35.96-38.02 g·cm-2,中海拔草地为 43.27-45.12 g·cm-2,高海拔草地为40.36-42.13 g·cm-2,大致表现为中海拔>高海拔>低海拔,不同海拔草地BGB差异均显著(P<0.05);低海拔草地根冠比(R/S)为0.64-0.65,中海拔草地为 0.75-0.78,高海拔草地为 0.71-0.73,大致表现为中海拔>高海拔>低海拔,不同海拔草地R/S差异均显著(P<0.05)。
2.2 不同海拔山地草地物种丰富度、多样性及均匀性
植物群落丰富度分析结果表明(表 3),研究区山地群落丰富度指数随海拔升高呈先增加后降低趋势。低海拔草地丰富度指数为 7.25-8.26,中海拔草地为9.36-12.35,高海拔草地为8.56-9.17,大致表现为中海拔>高海拔>低海拔,高海拔和低海拔丰富度指数差异不显著(P<0.05);低海拔草地均匀度指数为 0.79-0.82,中海拔草地为 0.86-0.87,高海拔草地为 0.83-0.84,大致表现为中海拔>高海拔>低海拔,高海拔和低海拔均匀度指数差异不显著(P<0.05);低海拔草地优势度指数为2.15-2.17,中海拔草地为 1.98-1.99,高海拔草地为2.01-2.04,大致表现为低海拔>高海拔>中海拔,高海拔和中海拔优势度指数差异不显著(P<0.05);低海拔草地多样性指数为 0.76-0.79,中海拔草地为 0.87-0.91,高海拔草地为 0.82-0.83,大致表现为中海拔>高海拔>低海拔,不同海拔草地多样性指数差异均显著(P<0.05)。
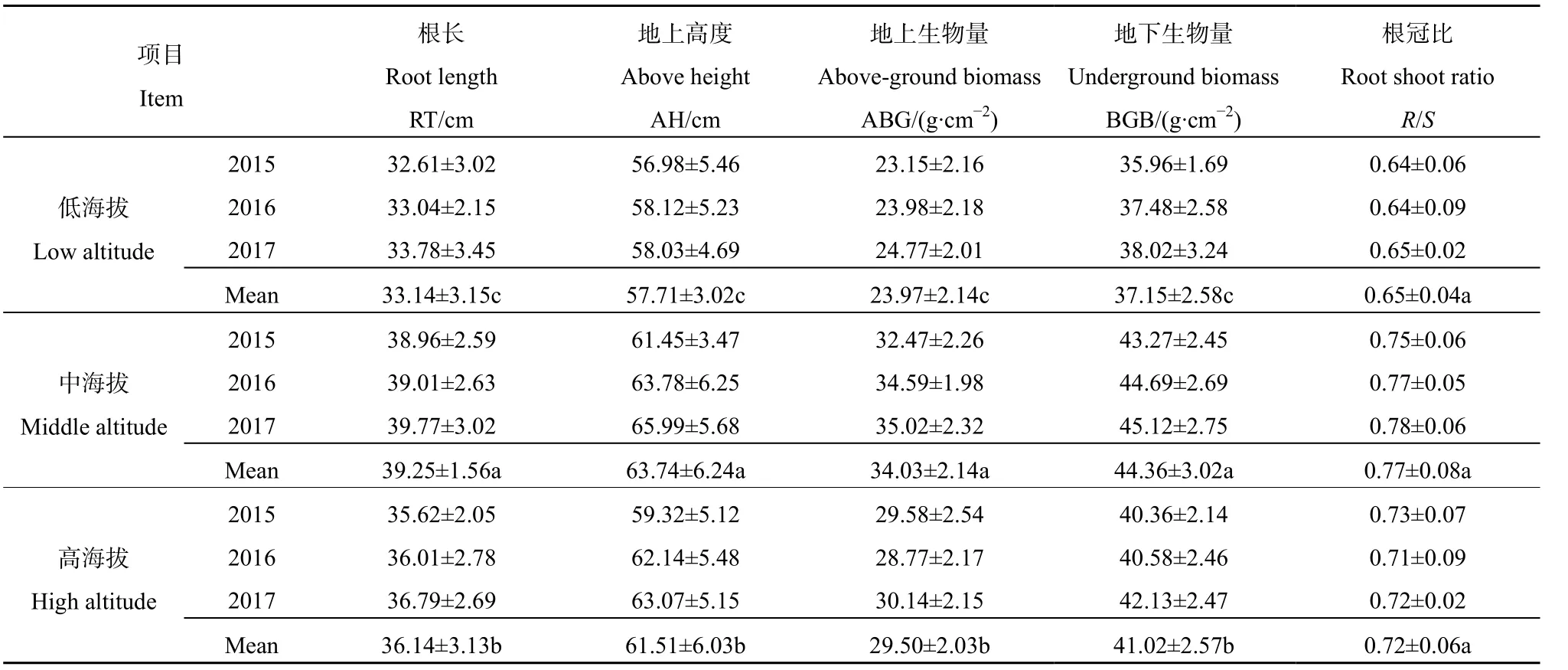
表2 不同海拔山地草地地上与地下生物量Table 2 Above and underground biomass of grassland at different elevations
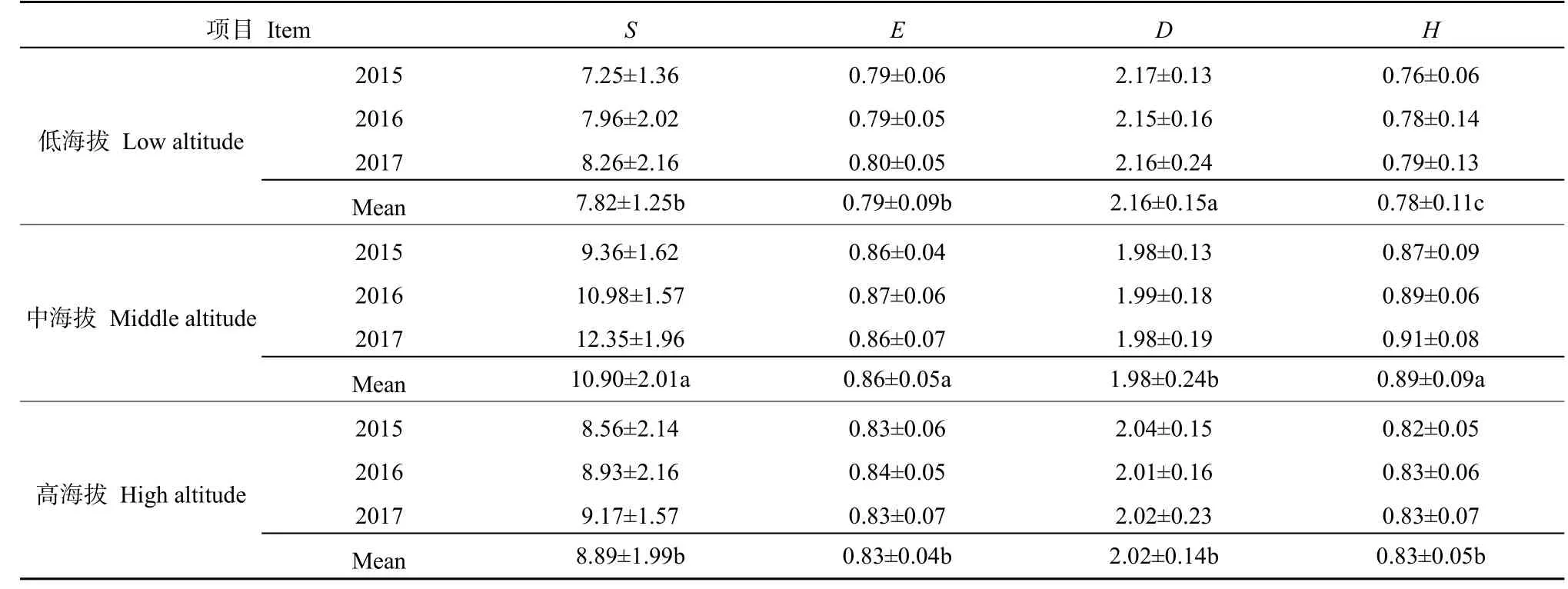
表3 不同海拔草地植被群落物种丰富度、多样性及均匀性Table 3 Plant community species richness, diversity and evenness of grassland at different elevations
2.3 不同海拔山地草地土壤理化性质特征
由表4可知,低海拔草地土壤pH为0.76-0.79,中海拔草地为 0.87-0.91,高海拔草地为 0.82-0.83,大致表现为中海拔>高海拔>低海拔,不同海拔草地土壤 pH差异均显著(P<0.05)。低海拔草地土壤全碳质量分数为 12.36-13.25 g·kg-1,中海拔草地为15.48-15.67 g·kg-1,高海拔草地为13.56-13.89 g·kg-1,大致表现为中海拔>高海拔>低海拔,不同海拔草地土壤全碳质量分数差异均显著(P<0.05)。低海拔草地土壤全氮质量分数为 1.16-1.17 g·kg-1,中海拔草地为 1.28-1.30 g·kg-1,高海拔草地为1.22-1.23 g·kg-1,大致表现为中海拔>高海拔>低海拔,不同海拔草地土壤全氮质量分数差异均显著(P<0.05)。低海拔草地土壤全磷质量分数为0.98-0.99 g·kg-1,中海拔草地为1.00-1.01 g·kg-1,高海拔草地为 0.98-1.01 g·kg-1之间,大致表现为中海拔>高海拔>低海拔,不同海拔草地土壤全磷质量分数差异均不显著(P>0.05)。草地土壤全钾、碱解氮和速效磷均表现为中海拔>高海拔>低海拔,差异均达显著水平(P<0.05)。
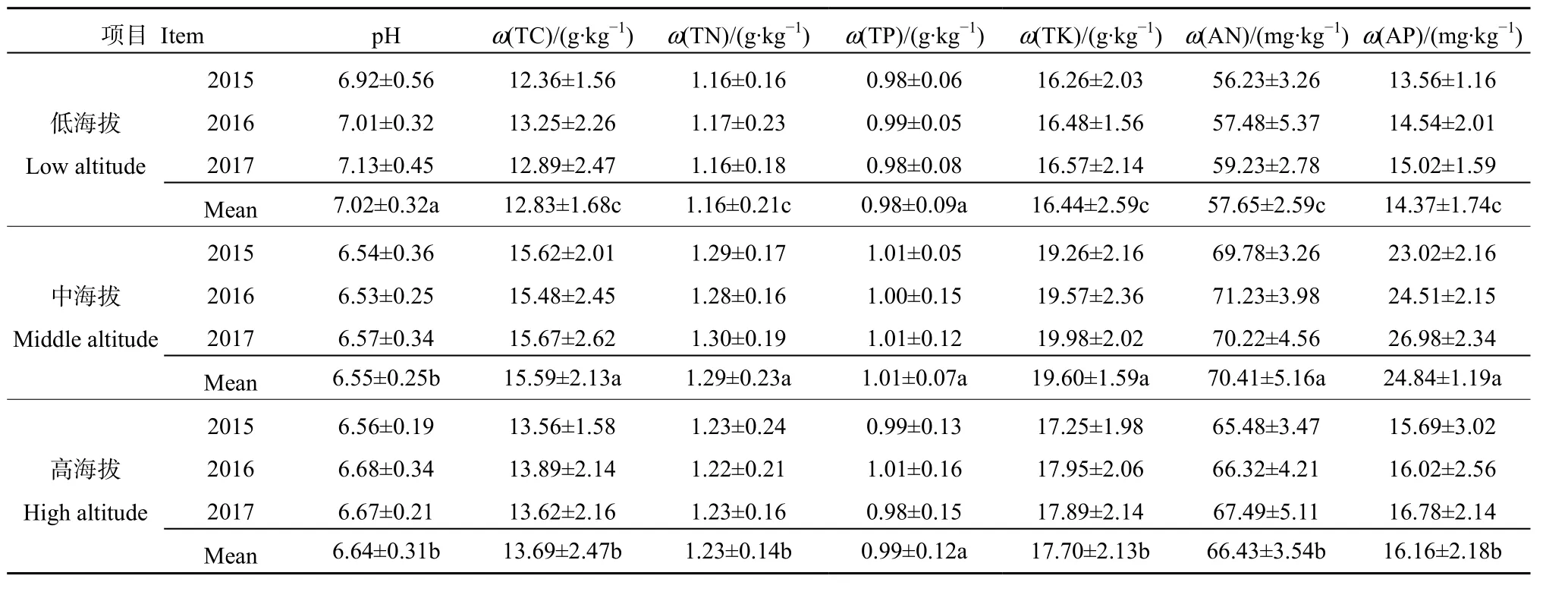
表4 不同海拔山地草地土壤理化特征Table 4 Soil physical and chemical properties of grassland at different elevations
2.4 不同海拔山地草地生物量与土壤理化特征关系
土壤理化特征对植被的生长起着重要的作用,为探讨山地草地生物量与土壤理化特征关系,对生物量与土壤理化特征进行相关性分析(表5)。结果表明,植被多样性各指标和生物量均与pH之间存在负相关关系,与土壤理化特征等均呈正相关关系。物种丰富度指数(S)和多样性指数(H)与土壤理化特征的相关系数绝对值均高于优势度指数(D)和均匀度指数(E),说明了土壤理化特征对植被优势度指数和均匀度指数的影响作用较小。由此可知,土土壤理化特征与植被多样性密切相关,其中土壤pH对植被多样性贡献为负,土壤理化特征对植被多样性贡献为正。土壤全碳和全氮与根长和生物量呈显著正相关(P<0.05),表明全碳和全氮是影响根长和生物量的主要养分指标。
2.5 土壤理化特征与植被群落的典范相关分析
将喀斯特山地草地的影响因子分成3组变量,其中,土壤理化性质指标(pH值、全碳、全氮、全磷、全钾、速效磷、速效氮、速效钾)构成第 1组变量,生物量(地上和地下生物量)和植被多样性分别构成第 2、3组变量。采用典范相关分析研究土壤理化特征与植被群落之间的关系,由表6可知,生物量对多样性的贡献最大。
环境因子在植被群落分布中起着重要的作用,水热及地形地貌等都会对植被的分布产生直接作用,在植被群落研究中常采用冗余分析(RDA)来探究二者之间的关系。其中的响应变量是植物群落多样性,解释变量为土壤理化性质和生物量。由图2可知,前两个排序轴的累积解释率达到了80%,而第一个因子解释率达到了62%,且通过了显著性统计检验,可认为是该部分环境因子对植被产生显著的制约作用。此外,通过研究发现,除土壤 pH值外,植被多样性越丰富,土壤理化性质越好,二者之间存在显著的正相关关系;在土壤理化性质不断改善的情况下,植物丰富度呈现明显的上升,其中相关性最大的是全碳及全氮,这是植物群落分布的最主要影响因子,与相关性分析结果一致。
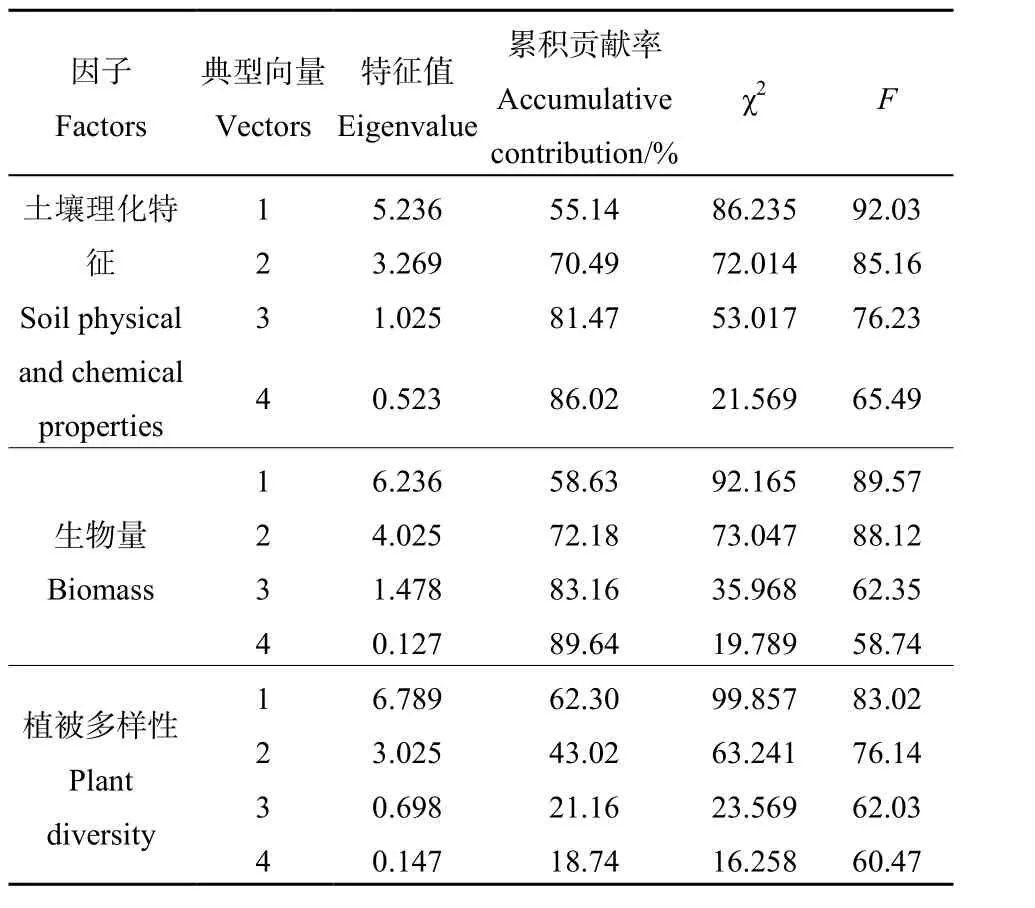
表6 土壤理化特征与植被群落的典范相关分析Table 6 Canonical correlation analysis between biomass and soil physical and chemical properties
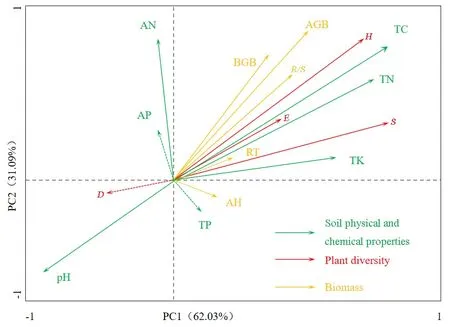
图2 土壤理化特征与植被群落的典范RDA排序图Fig. 2 RDA sequence diagram between biomass and soil physical and chemical properties
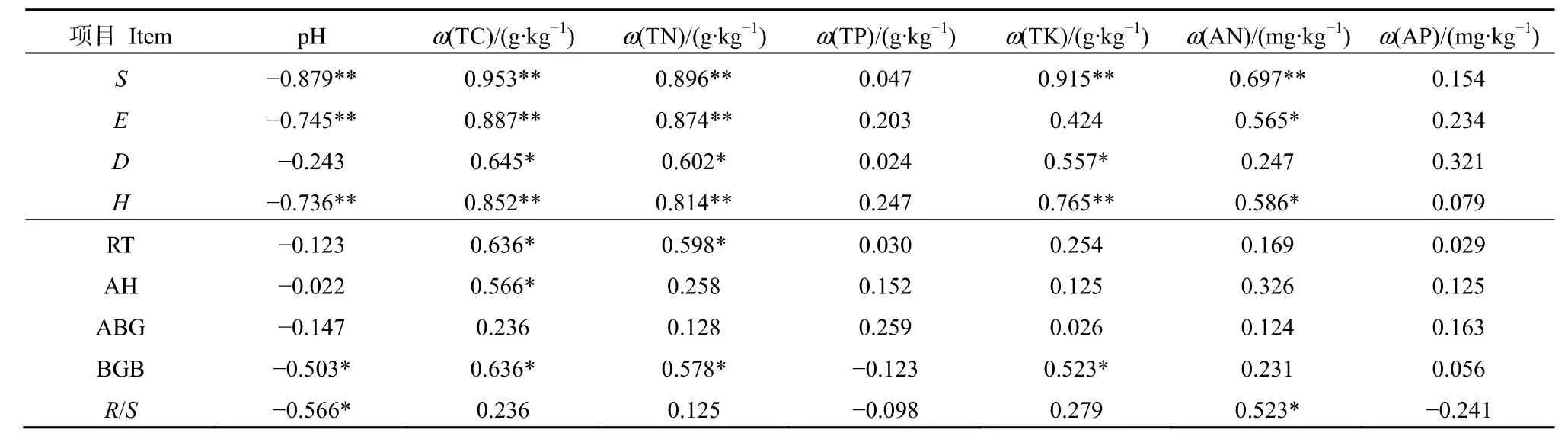
表5 不同海拔山地草地生物量与土壤理化特性关系Table 5 Relationship between biomass and soil physical and chemical properties at different elevations
3 讨论
群落学特征是草地生产、生态稳定性调控研究的基础。只有了解草地的群落组成、结构和功能,才能更好地实现草地的高效培育和合理利用,使草地的生产、生态功能得以全面发挥(John et al.,2007;Chen et al.,2018)。植物群落物种多样性是生物及其与环境形成的生态复合体以及与此相关的各种生态过程的总和,不仅能够度量群落的组成结构和功能复杂性,而且也能指示环境状况(Yadeta et al.,2018)。本研究表明,不同海拔草地根长、高度、地上和地下生物量在 2015-2017年呈现增加的趋势,但增幅不明显,3年平均值基本表现为中海拔>低海拔>高海拔,并且不同海拔差异均显著(P<0.05),与前人的研究结果一致(Chick et al.,2018;Busch et al.,2018)。有研究认为群落的生产力水平为中等水平时其生物多样性最大,生产力和多样性指数呈钟型曲线关系,即多样性在低水平时随生产力的增加而增加,在达到足够高的生产力时反而下降(Shrestha et al.,2018);同时,也有研究认为,在中海拔梯度植物群落中,土壤养分含量相对较低,这在一定程度上抑制了某些植物的充分生长,表现为生产力下降,然而这种生产力的降低因物种间或功能群间的相互协同而得到补偿,即某一植物种或植物功能群生物量的减少部分由另外一些植物种或植物功能群生物量的增加部分所补偿,故生产力能够维持在一定水平上(Yadeta et al.,2018)。因此,特定资源生产力水平下草地群落固有的生物多样性,是保持草地稳定和健康发展的基础(Ren et al.,2018;Zhang et al.,2018;De et al.,2018)。
土壤作为生态系统中生物与环境相互作用的基质,贮存着大量的碳、氮、磷等营养物质,土壤养分对植物生长具有至关重要的作用,直接影响着植物群落的种类组成与生活性特征,决定着生态系统的结构和功能(Verdú et al.,2018)。植物在养分丰富的局部环境中能选择性地改变其根系的生长,从而增加养分的吸收。有研究表明,土壤中养分含量的高低直接影响着群落的生产力,土壤养分越丰富,群落生产力越高(Verdú et al.,2018;Zhang et al.,2018)。本研究中,不同海拔草地土壤全碳、全氮、全钾、碱解氮和速效磷均表现为中海拔>高海拔>低海拔,差异均达显著水平(P<0.05);土壤pH值与之呈相反的变化趋势,而土壤全磷在不同海拔之间差异不显著(P>0.05);随着海拔的增加,土壤水分含量逐渐增加,植物凋落物和有机质含量亦逐渐增加,土壤微生物呼吸加强,对土壤养分的利用效率提高,海拔越高,土壤的有机质越丰富,为土壤微生物生长提供更多碳源和可利用养分,促进土壤微生物的大量繁殖及活动,导致土壤微生物对有机质的利用效率和矿化速率提高,从而改善土壤理化性质(Pingree et al.,2018;Zhang et al.,2018)。然而,本研究中高海拔草地土壤理化特征和植被多样性植被有所下降,主要是由于在高海拔温度下降显著,从低海拔的水分限制到高海拔的温度限制,土壤理化特征、生物量和植物群落生物量随海拔呈先增加后降低趋势。磷含量的变化与海拔没有必然的联系,主要受到成土母质的影响(Shrestha et al.,2018)。相关性分析表明:植被多样性各指标和生物量均与 pH之间存在负相关关系,与其他土壤理化特征等均呈正相关关系,由此可知在植被分布中,pH对其起着一定的制约作用,而全碳和全氮是影响根长和生物量的主要土壤因子。
4 结论
通过对不同海拔喀斯特山地草地群落多样性和土壤理化性质2015-2017年连续3年的研究发现,不同海拔山地草地根长、高度、地上、地下生物量、土壤理化指标(除了全磷)均呈增加的趋势,而土壤pH值呈相反的变化趋势。3年间,山地草地根长、高度、地上、地下生物量、土壤理化特征(除了全磷)增幅不明显,但不同海拔差异均显著(P<0.05),说明海拔对山地草地根长、高度、地上、地下生物量、土壤理化特征(除了全磷)的影响较大。植被丰富度指数、均匀度指数和多样性指数随海拔的增加呈先增加后降低的趋势,在中海拔表现最高,说明中海拔有利于植被多样性的提高。土壤全碳和全氮与植被多样性呈显著正相关,表明全碳和全氮是影响植被多样性的主要土壤环境因子。
