极端干旱对荒漠草原群落物种多样性和地上生物量碳氮的影响
2019-05-31张蕊赵学勇王少昆左小安王瑞雄
张蕊 ,赵学勇,王少昆,左小安,王瑞雄
1. 中国科学院西北生态环境资源研究院,乌拉特荒漠草原研究站/奈曼沙漠化研究站,甘肃 兰州 730000;2. 中国科学院大学,北京 100049
荒漠生态系统是全球最大的陆地生态系统之一,约占陆地总面积的41%,为全球37%的人口提供生计资料和生活环境(Reynolds et al.,2007)。荒漠化草原是典型草原向荒漠过渡的一个脆弱生态带,易受气候变化和人类活动的共同影响(周培等,2011),特别是极端降水事件的影响。降水是干旱区荒漠生态系统的重要水分来源(张腊梅等,2014)。荒漠区降水时空分布是决定植物发芽、定植、生长、群落物种组成和生物量(Dasci et al.,2010)的极其重要的因素(Cheng et al.,2006)。中国荒漠草原总面积达1.9×105hm2,占全国草原总面积的4.82%(曲浩等,2014)。内蒙古荒漠草原是世界草原植被中特有的植被类型,是旱生性最强的草原生态系统(中国科学院内蒙宁夏综合考察队,1985),在生态地理条件、群落结构和功能方面都具有独特性。位于内蒙古自治区西北部的乌拉特荒漠草原属于荒漠植物成分强烈侵入草原的植被类型,属于半干旱区与干旱区的边缘地带,处于荒漠与典型草原的过渡带上(张蕊等,2019)。
物种多样性既能度量群落的结构组成和功能复杂性,也能指示环境承载力的大小(左小安等,2007)。因此,物种多样性作为生物多样性的主要研究层次备受关注。群落物种多样性与植被生产力是衡量草地生态系统生态和生产功能的重要指标,也是维持草地生态系统可持续稳定发展的基础(Tilman et al.,1996),它们可以体现群落的结构、群落组织水平、发展阶段、稳定性程度和生境差异。碳储量是生态系统碳库的重要组成成分,在全球碳循环中发挥着重要的作用(林婉奇等,2019)。植被碳储量对生态系统碳平衡具有重要调节作用,过去100年全球植被碳储量整体上呈增加趋势(孙晓芳等,2013)。生物量碳密度是生态系统表征碳截存能力的重要功能特征之一(张蕊等,2018)。
在全球气候变化大背景下,亟需了解干旱半干旱地区草原生态系统的结构、功能和过程如何响应极端气候事件,这对于我们认知自然生态系统的演变规律,预防和降低未来极端气候事件对生态环境和经济发展造成的损失,均具有重要意义(张彬等,2014)。据预测,由于极端气候事件的发生,陆地生态系统的干旱程度和频率将会增加(IPCC,2013;Luo et al.,2018),并对其产生重大影响,特别是对物种多样性和生产力受降水严重限制的干旱和半干旱生态系统(Abramsky et al.,1984)。干旱通过改变物种多样性,降低草地生产力(韩彬等,2006;孙小丽等,2015)和改变植物碳、氮的质量分数(Luo et al.,2018)而影响植被碳密度,最终影响生态系统的碳截存能力。然而,目前关于极端干旱对植物碳、氮影响的研究(Luo et al.,2018)和不同极端干旱类型对荒漠草原生态系统影响的研究较少。
基于此,本研究选取乌拉特荒漠草原沙生针茅群落为研究对象,分析极端干旱处理下沙生针茅(Stipa glareosa)群落物种多样性和生产力变化特征,揭示干旱对地上生物量碳、氮的影响规律,进而阐明不同极端干旱处理对荒漠草原生态系统碳密度的影响机理,为深刻认识未来气候变化下荒漠草原生态系统结构和功能特征提供科学数据和理论依据。
1 研究地概况与研究方法
1.1 研究区自然概况
研究地位于蒙古高原的乌拉特后旗(105°14′-107°36′E,40°40′-42°22′N),海拔 1650 m,属典型的大陆性干旱气候,热量丰富,其中≥10 ℃的年积温为2000-3000 ℃,年均气温5.7 ℃,年均风速5.3 m·s-1,一年内10 m·s-1以上风速日数达52 d,年均降雨量为 145 mm(1987-2017),年蒸发量超过降雨量的10倍;湿润系数为0.15-0.30,无霜期130 d左右。该区植被类型以荒漠草原和荒漠灌丛为主,土壤以棕钙土和灰棕漠土为主(赵学勇等,2014)。
研究样地位于中国科学院西北生态环境资源研究院乌拉特荒漠草原生态系统研究站围封的荒漠草原长期试验观测场内。该试验场从 2010年起开始围封,从南到北可划分为沙生针茅群落、芨芨草(Achnatherum splendens)群落和红砂(Reaumuria songarica)群落三大类型。围封的试验场能代表乌拉特荒漠草原的主要植被类型(曲浩等,2014)。
1.2 试验设计与调查方法
在沙生针茅群落典型地段设置极端干旱处理的观测样地,极端干旱试验装置由支撑架、截雨棚组成。极端干旱处理是通过支撑架上安装的截雨棚(聚碳酸酯塑料)截雨实现的。该试验平台建于2015年5月,共12套极端干旱处理试验装置,极端干旱处理有生长季 5-8月截雨 66%(-66%),6-7月干旱60 d(-100%)和对照(CK)。为避免处理间的相互干扰,每个样方周围留出2 m的缓冲带。每个处理6个重复,共18个6 m×6 m的样方。
于2017年6月下旬和8月中下旬各调查一次每个样方内的植物种类、盖度(针刺法,1 m×1 m样方框分割成100个0.1 m×0.1 m的小格子)和高度(用卷尺测量植株的自然高度,每种植物测3-5株),每个样方内各调查4个1 m×1 m的小样方,用于分析计算物种多样性。8月调查完植物种类、盖度和高度后,于每个样方内设置1个1 m×1 m的小样方,将所有植物齐地面刈割,并收获小样方内的凋落物。每个样方内的现存生物量和凋落物分开装入信封,带回实验室,在 65 ℃条件下烘干至恒重,用于计算地上生物量。称重后样品粉碎,用元素分析仪(Costech ECS4010,Italy)测定样品中碳、氮的质量分数。
1.3 物种多样性指数和生物量碳、氮密度计算公式
物种多样性指数采用物种丰富度(物种数S)指数、Shannon-Wiener指数、Simpson优势集中性指数和Pielou均匀度指数(乔文静等,2018)表征。群落中物种优势程度根据物种重要值确定,物种重要值体现了物种在群落中的优势程度(葛兆轩等,2017)。本文用植物种的相对盖度(CR)、相对高度(HR)和相对频度(FR)来表示其重要值(IV)。具体公式如下:

物种丰富度指数(Species richness):

多样性指数(Shannon-Wiener指数):

Simpson优势集中性指数:

均匀度指数(Pielou均匀度指数):

式中,S为样方种出现的物种总数;Pi为第 i种的相对重要值。

式中,BCD、BND分别表示生物量碳和生物量氮密度,g·m-2;B表示单位面积上的生物量,g·m-2;CC、NC 分别表示生物量碳的质量分数和生物量氮的质量分数,g·kg-1(常天军等,2007)。
1.4 数据分析
运用 Excel整理数据,SPSS进行统计分析,SigmaPlot 12.5进行绘图。不同极端干旱处理间物种多样性,生物量,生物量碳、氮的质量分数和生物量碳、氮密度采用单因素方差分析(ANOVA)法进行差异性比较。差异显著性检验采用 Duncan's法,显著水平为0.05。
2 结果
2.1 研究区降雨量特征
研究区1987-2017年年均降雨量为145 mm,生长季(5-9月)平均降雨量为124 mm,占年均降雨量的85%(图1)。其中年降雨量和生长季降雨量最低年份都是2001年,分别为63 mm和50 mm;年降雨量最高年份是2003年(235 mm),生长季降雨量最高年份是1994年(220 mm)。在1987-2017这30年内,最低年降雨量是年均降雨量的44%,最高年降雨量是年均降雨量的 162%。因此,该试验极端干旱处理我们选择了生长季 5-8月截雨66%和6-7月干旱60 d。

图1 研究区平均月降雨量(1987-2017)Fig. 1 Average monthly precipitation in study area (1987-2017)
研究区2017年降雨量为154 mm,其中生长季5-8月的降雨量为122 mm。5-8月截雨66%和6-7月干旱60 d两种极端干旱处理下样地在生长季5-8月实际获得的降雨量分别为41 mm和76 mm(图2)。
2.2 群落物种组成及重要值对极端干旱的响应

图2 极端干旱处理下实际月降雨量(2017)Fig. 2 Monthly precipitation of different treatments in study area (2017)
18个研究样方内共调查到16种植物,其中半灌木3种:戈壁天门冬(Asparagus gobicus)、猫头刺(Oxytropis aciphylla)、蓍状亚菊(Ajania achillodes),半灌木状草本 1种薄叶燥原荠(Ptilotrichum tenuiflium),其他为草本。经过3年极端干旱处理后,荒漠草原沙生针茅群落物种组成及其重要值发生了改变,但不同极端干旱处理下,优势物种未发生变化。由表1中的重要值可知,群落优势物种为沙生针茅和骆驼蓬(Peganum harmala)。在6月,与对照相比,-66%极端干旱处理下没有出现的物种有薄叶燥原荠,新增的物种有沙茴香(Ferula bungeana)、冬青叶兔唇花(Lagochilus ilicifolius)和蒙古韭(Allium mongolicum);-100%极端干旱处理下没有出现的物种有冷蒿(Artemisia frigida)和白花黄芪(Astragalus leucocephalus),新增的物种有沙茴香、冬青叶兔唇花和五星蒿(Bassia dasyphylla)。在8月,与对照相比,-66%极端干旱处理下没有出现的物种有蓍状亚菊,新增的物种有冬青叶兔唇花;-100%极端干旱处理下没有出现的物种有白花黄芪和蓍状亚菊,新增的物种有冬青叶兔唇花。这表明冬青叶兔唇花只在极端干旱处理下出现。对照下,8月调查到而6月没有调查到的物种有沙茴香、蒙古韭、五星蒿、碱韭(Allium polyrhizum)、蓍状亚菊和薄翅猪毛菜(Salsola pellucida),但是薄叶燥原荠只在6月调查到(表1)。
2.3 物种多样性对极端干旱的响应
Shannon-Wiener指数、Pielou均匀度指数、Simpson优势集中性指数和物种丰富度指数都表现为8月高于6月。6月不同处理间4种物种多样性指数基本保持一致,而 8月两种极端干旱处理都降低了Shannon-Wiener指数和物种丰富度指数,仅-66%处理降低了Simpson优势集中性指数(图3)。
2.4 地上生物量对极端干旱的响应
与对照相比,两种极端干旱处理显著降低了地上现存生物量,但只有-66%处理显著降低了凋落物生物量(图 4)。对照的地上现存生物量为 29.6 g·m-2,凋落物生物量为 15.2 g·m-2。-66%和-100%极端干旱处理的现存生物量较对照分别减少65%、和56%;-66%和-100%极端干旱处理的凋落物生物
量较对照分别减少 61%、20%。在对照、-66%和-100%处理下,地上现存生物量与凋落物生物量的比值依次为1.9、1.7和1.1。

表1 极端干旱处理下荒漠草原群落物种组成及其重要值Table 1 Species composition and importance value under different extreme drought treatments
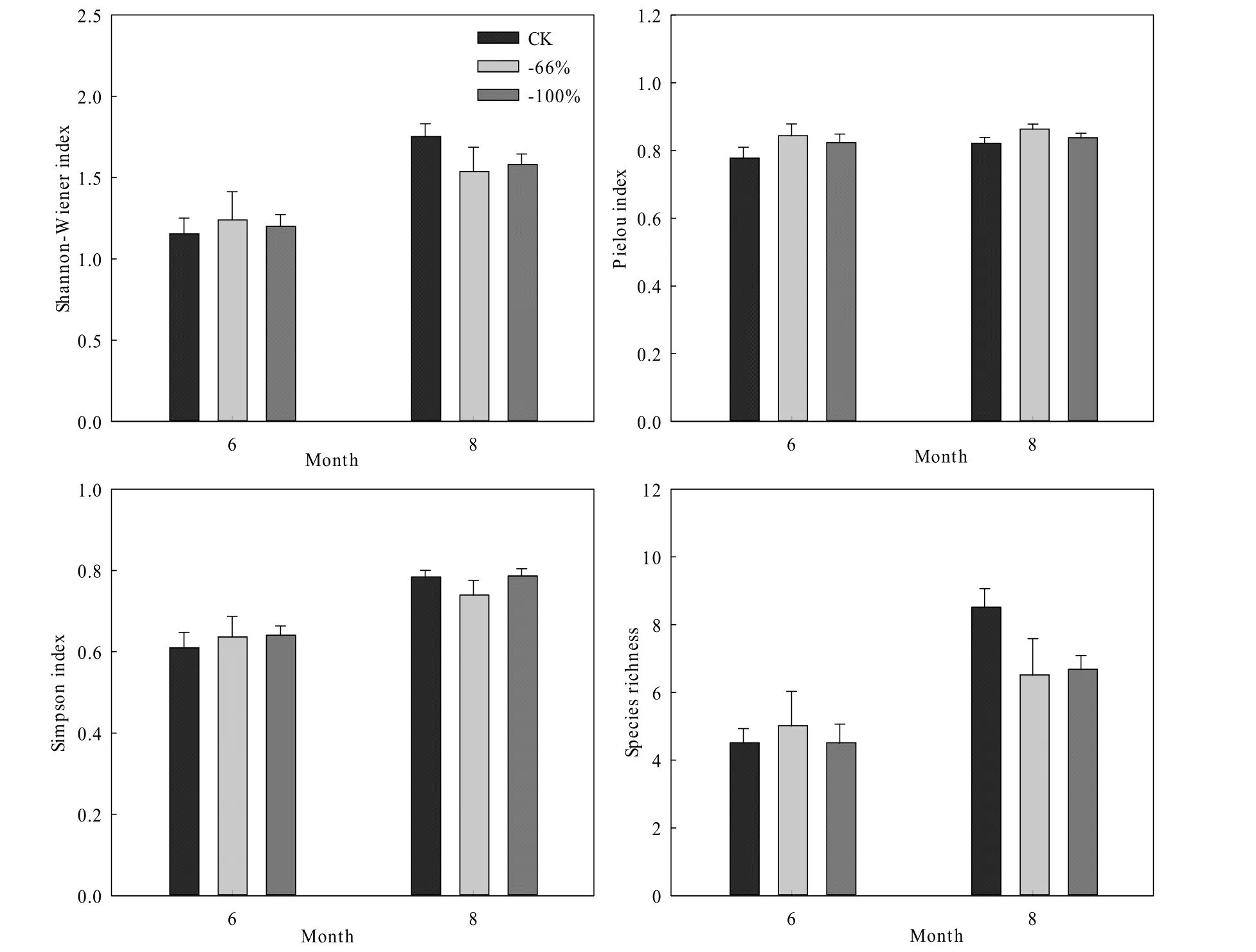
图3 物种多样性对极端干旱处理的响应Fig. 3 Response of community species diversity to extreme drought treatments

图4 极端干旱处理对地上生物量的影响Fig. 4 Effect of extreme drought treatments on aboveground biomass
2.5 地上生物量碳氮对极端干旱的响应
现存生物量碳的质量分数范围为 416.3-424.6 g·kg-1,凋落物生物量碳的质量分数为 350.3-377.1 g·kg-1。不同极端干旱处理对现存生物量和凋落物生物量碳的质量分数的影响无显著差异(图5A)。现存生物量氮的质量分数为27.3-34.5 g·kg-1,凋落物生物量氮的质量分数为10.1-10.9 g·kg-1。-100%极端干旱处理下,现存生物量氮的质量分数显著高于对照和-66%极端干旱处理的。但凋落物生物量氮的质量分数在不同处理间无显著差异(图5B)。
现存生物量碳密度范围为4.3-12.6 g·m-2,凋落物生物量碳密度为2.3-5.4 g·m-2。与对照相比,两种极端干旱处理显著降低了现存生物量碳密度;而只有-66%极端干旱处理显著降低了凋落物生物量碳密度(图6A)。现存生物量氮密度范围为0.27-0.81 g·m-2,凋落物生物量氮密度为 0.07-0.15 g·m-2。与对照相比,两种极端干旱处理显著降低了现存生物量氮密度;而只有-66%极端干旱处理显著降低了凋落物生物量氮密度(图6B)。
3 讨论
3.1 极端干旱条件下物种组成和物种多样性的变化
极端干旱处理改变了荒漠草原沙生针茅群落的物种组成。6月生长季5-8月截雨66%和6-7月干旱 60 d两种极端干旱处理的物种数较对照分别增加了2种和1种。而8月6-7月干旱60天干旱处理的物种数较对照减少了1种。在6月,极端干旱处理下物种数比对照多的可能原因是,土壤潜在种子库中原本休眠的部分种子被极端干旱处理打破了休眠,从而正常发芽生长。例如冬青叶兔唇花,对照下6月、8月都没有调查到,而两种极端干旱处理下都出现了。具有耐旱和避旱机制的植物不会受干旱条件的影响,甚至会生长得更好,然而缺乏耐旱机制的植物将会减少,甚至大量死亡(Munson et al.,2012;曾勇,2015)。这点解释了为什么在6月极端干旱处理下物种数较对照多。薄叶燥原荠只在6月调查到,这可能是其在8月底之前就完成了生活史。碱韭、蓍状亚菊和薄翅猪毛菜只在8月调查到,说明这些植物种在荒漠草原生态系统中仅在生育期晚或在生长季中后期降雨后才能发芽生长。

图5 极端干旱处理对地上生物量碳(A)、氮(B)质量分数的影响Fig. 5 Effect of extreme drought treatments on aboveground biomass carbon (A) and nitrogen (B) mass fraction
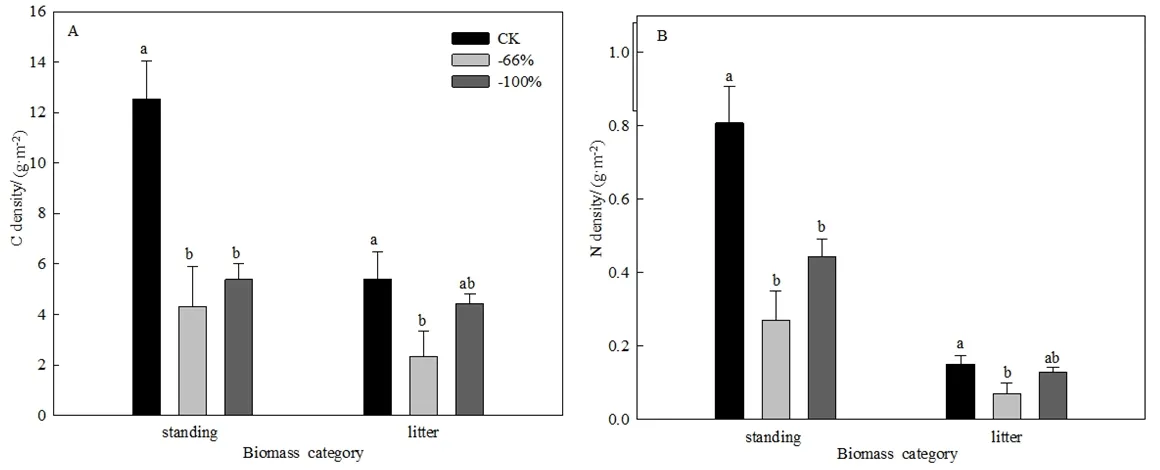
图6 极端干旱处理对地上生物量碳(A)、氮(B)密度的影响Fig. 6 Effect of extreme drought treatments on aboveground biomass carbon (A) and nitrogen (B) density
物种多样性可以反映草地生态系统的基本状况,多样性增加有助于系统稳定健康发展。整体而言,4种物种多样性指数都呈现6月低于8月,这是因为生长季前期本试验所在区域内降雨极少,抑制了植物的发芽生长,生长季中期降雨后,有些植物才开始发芽定植生长,例如碱韭、蓍状亚菊和薄翅猪毛菜。土壤水分影响物种多样性,土壤水分对植物群落的影响首先是减少了植物种的丰富度,进而威胁到群落规模,最终导致植被盖度下降(韩路,2014)。8月极端干旱处理降低了 Shannon-Wiener指数和物种丰富度指数,而Pielou均匀度指数在不同极端干旱处理下基本保持不变。研究表明,物种丰富度指数、Shannon-Wiener指数、Simpson指数均与年降水量呈显著的正相关关系(孙小丽等,2015),但年降水量对 Pielou均匀度指数的影响不显著(杨崇曜等,2017)。因此,极端干旱会降低土壤水分,影响植物生长发育,最终影响群落物种多样性。
3.2 极端干旱条件下植物生物量及其碳氮的变化
研究表明,干旱、半干旱区植物群落地上净初级生产力与平均年降水量呈显著正相关关系(Sala et al.,1992;Knapp et al.,2001;孙小丽等,2015)。因此,平均年降水量被认为是草地生产力一个最关键的驱动因子(Chou et al.,2010;张彬等,2014)。降水量每增加1 mm,青海曲麻莱县天然草地产草量增加 24 kg·hm-2(吕晓英,2003;罗冬,2016)。生长季5-8月截雨66%和6-7月干旱60 d两种极端干旱处理都使地上现存生物量较对照减少 50%以上。与对照相比,两种极端干旱处理也减少了凋落物生物量。其中,生长季 5-8月截雨66%使凋落物生物量降得更多。这表明极端干旱处理降低了地上生物量的积累。这主要是因为极端干旱事件导致植物水分胁迫(张彬等,2014),水分胁迫影响植物生长发育(Bernal et al.,2011;牟成香等,2013),最终改变生物量的积累。极端干旱处理降低了地上现存生物量与凋落物生物量的比值,这也说明极端干旱改变了荒漠草原沙生针茅群落生物量的积累,干旱使地上现存生物量相对减少,而使凋落物生物量相对增加。有研究指出,同一物种的生物量在不同功能群组成样地之间存在明显差异,竞争关系的改变是影响物种生物量变化的一个重要因子(王江,2006)。因此,极端干旱处理下,物种组成的改变可能也是地上生物量减少的一个重要原因。
极端干旱处理对地上生物量碳的质量分数无显著影响,但6-7月干旱60 d的干旱处理显著提高了地上现存生物量的氮的质量分数。植物冠层养分含量与干旱呈正相关关系,即降雨量减少和干旱处理提高了植物冠层氮的质量分数(Luo et al.,2015)。这表明极端干旱有助于提高植物氮的质量分数。虽然极端干旱处理对地上生物量碳的质量分数无显著影响,但两种极端干旱处理使地上现存生物量的碳密度较对照降低了50%以上。而且生长季5-8月截雨 66%干旱处理也显著降低了凋落物生物量的碳密度。极端干旱处理对生物量氮密度有相似的影响。这主要是由于极端干旱处理改变了生物量引起的。
极端干旱通过影响荒漠草原群落物种组成和生长发育进而影响植物群落结构和功能,最终影响生态系统碳截存和碳收支过程。本研究通过分析荒漠草原群落物种多样性、生物量和生物量碳、氮密度对极端干旱的响应,为进一步深入分析干旱半干旱区植被对极端降雨事件的响应机制奠定基础。
4 结论
极端干旱改变了荒漠草原沙生针茅群落的物种组成。在8月份,生长季5-8月截雨66%和6-7月干旱60 d两种极端干旱处理都降低了群落的Shannon-Wiener指数和物种丰富度指数。两种极端干旱处理使地上现存生物量较对照降低了 50%以上。生长季5-8月截雨66%干旱处理使凋落物生物量较对照降低了61%,同时显著降低了凋落物生物量的碳氮密度,两种极端干旱处理使地上现存生物量的碳氮密度较对照降低了50%以上。这表明极端干旱不仅改变了荒漠草原群落的物种组成,而且改变了地上生物量,影响了植被碳氮密度,从而减弱荒漠草原植物群落地上部分的碳氮汇功能。
